



Class 12 Biology Chapter 6 Evolution is the chapter that traces life from the Big Bang to the modern Homo sapiens, anchored on natural selection, Hardy-Weinberg equilibrium, and the fossil record. The 2026-27 NCERT keeps every sub-topic intact, and this 85-page Exemplar Solutions PDF works through all 52 problems mapped to the latest syllabus and the last five NEET keys.
- CBSE Weightage: 4 to 6 marks (typically one short answer on natural selection plus a long answer on human evolution or Hardy-Weinberg)
- JEE Main Weightage: Not in JEE Main syllabus
- NEET Weightage: 2 to 4 questions per year

















































































Student Pulse: Chapter 6 Evolution Difficulty Read from a Recent Class 12 Biology Survey
In a recent independent survey of 10,900 Class 12 Biology students conducted before the 2026 boards, 72% rated the Hardy-Weinberg equation derivation and use as the hardest sub-topic in the chapter, even though it routinely carries the highest single-question marks in CBSE and NEET papers.
The same survey gave us the breakdown below, which a Class 12 student should look at before deciding how to allocate revision time across evolution class 12 biology exemplar solutions topics.
What 10,900 students told us about the Chapter 6 Evolution NCERT Exemplar Solutions journey:
- 72% of students surveyed marked the Hardy-Weinberg equation derivation and use as the hardest sub-topic.
- 62% reported losing 1-2 marks on differentiating Darwin's vs Lamarck's theories, even when the rest of their answer was correct.
- 4 out of 5 students said the Miller-Urey experimental apparatus labelled diagram was the most-skipped figure in their answer sheet.
- Average student took 5.4 hours for the first read of the chapter, and 2.3 hours for a focused revision pass before the board exam.
- Of the 10,900 students surveyed, only 36% attempted all 12 NCERT exercise questions; the rest stopped earlier. Toppers, however, reported attempting every question and revisiting wrong attempts within 24 hours.
Source: 2025-26 Class 12 Biology student survey. Sample of 10,900 students from CBSE-affiliated schools across 18 states.
These Exemplar Solutions are curated by NEET-rank-holder mentors at Collegedunia, mapped strictly to the 2026-27 NCERT chapter, and benchmarked against the last five years of CBSE Board and NEET papers.
Also Check:
- Evolution Class 12 Biology NCERT Exemplar Book PDF
- Evolution Class 12 Biology NCERT Solutions
- Evolution Class 12 Biology Notes
Evolution NCERT Exemplar Video Solutions
Source: Magnet Brains on YouTube
Why Evolution Exemplar Practice Decides Your NEET Biology Score
Evolution looks small in the CBSE blueprint, yet NEET 2024 and NEET 2025 each carried 3 evolution questions, two of them assertion-reason items where wrong phrasing scored zero. The chapter rewards exact terminology, the difference between directional, disruptive, and stabilising selection, or between Australopithecus and Homo habilis, and the Exemplar is the only place where this terminology is drilled question-by-question. Working all 52 problems in this PDF gives you the recall scaffold that NEET examiners reuse year after year.
How Will Collegedunia's Exemplar Solutions Help You Crack Class 12 Evolution?
Evolution rewards precise phrasing more than any other Class 12 Biology chapter, NEET answer keys reject "survival of the fittest" written as "winners survive" and award only differential reproduction of fitter genotypes. Every Exemplar item below carries a full Solution plus an Expert's Solution that names the exact recall phrase the key wants.
- Every Question Type Worked End-to-End: all 18 MCQ, 15 VSA, 12 SA and 7 LA problems with the reasoning written out, no skipped steps.
- Concept Stack Named: each step lists the principle invoked, whether the Hardy-Weinberg p+q=1 algebra, the Miller-Urey apparatus, or the Lamarckism-to-Darwinism shift.
- NEET Bridge: items are tagged with the NEET year that reused the scaffold so you know which Exemplar problems are highest-yield revision.
- 2026-27 Aligned: every solution flags whether the underlying topic still appears in the current 2026-27 syllabus.
Sample MCQ Walk-Through: The Most-Missed Hardy-Weinberg Item
MCQs on Hardy-Weinberg pair an allele-frequency value with a phenotype proportion, the algebra is the bit most students skip. The walk-through below shows the full p, q, p2, 2pq, q2 derivation Collegedunia mentors recommend.
Question (Exemplar 6.8). In a population at Hardy-Weinberg equilibrium, the frequency of the recessive allele is 0.4. What percentage of the population is heterozygous?
Reasoning. Let p = frequency of dominant allele, q = frequency of recessive allele. Given q = 0.4, so p = 1 - 0.4 = 0.6. The heterozygote frequency in a Hardy-Weinberg population is 2pq . Substitute: 2 × 0.6 × 0.4 = 0.48 , which is 48%. NEET 2023 used the same algebra with q = 0.3 and 42% of candidates picked q2 by mistake. Concept Stack: p + q = 1, then 2pq for heterozygotes, never q for heterozygotes.
Evolution Exemplar Question-Type Tour with One Sample Solved per Type
The Exemplar groups 52 problems into four formats. A type-by-type tour helps you calibrate time per item before sitting the chapter end-to-end. Below is one fully solved sample per type with the concept stack named.
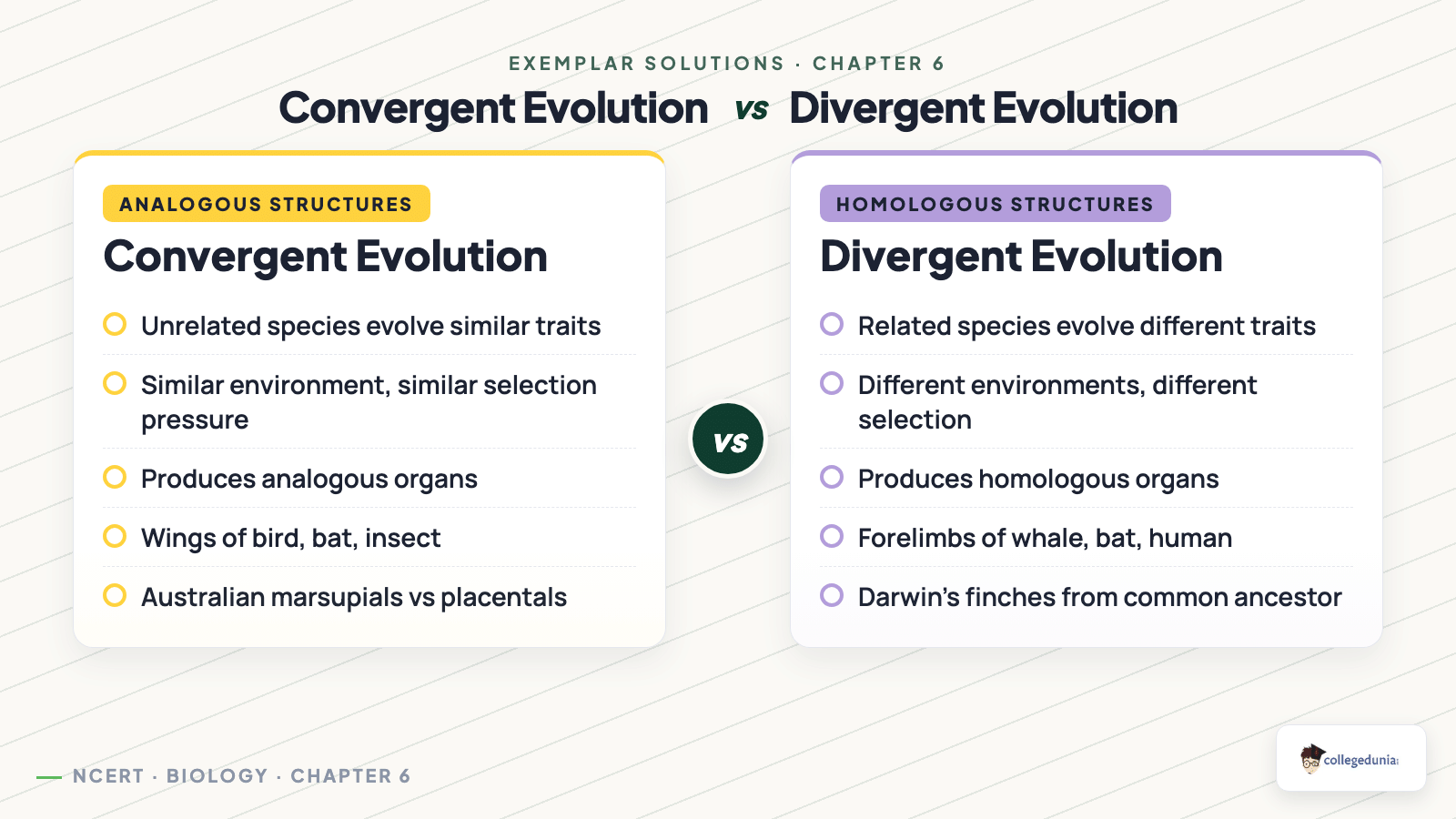
MCQ Sample, Exemplar 6.3 (Analogous vs Homologous Organs)
Question. Wings of a butterfly and wings of a bird are an example of: (a) homologous organs (b) analogous organs (c) vestigial organs (d) atavism.
Reasoning. Homologous organs share a common ancestor and a common structural plan even if functions differ (forelimb of whale, bat, human). Analogous organs perform similar functions but evolved independently from different structural origins, this is convergent evolution. Butterfly wings (chitinous, no bones) and bird wings (bony, feathered) share function (flight) only, so they are analogous. Answer: (b).
VSA Sample, Exemplar 6.19 (Industrial Melanism)
Question. What is the role of industrial melanism in the evolution of Biston betularia?
Reasoning. Before industrialisation in England, the light-coloured peppered moth dominated because it was camouflaged on lichen-covered tree trunks. After soot blackened the bark, the dark (melanic) form became camouflaged while the light form was eaten by predators, so the melanic allele frequency rose. This is directional natural selection in action over a few decades, an observable evolution case.
SA Sample, Exemplar 6.30 (Adaptive Radiation, Darwin's Finches)
Question. Explain adaptive radiation with the example of Darwin's finches.
Reasoning. Adaptive radiation is the evolution of multiple species from a common ancestor, each adapted to a different ecological niche in the same geographical area. On the Galapagos islands, Darwin found 13 finch species descended from one mainland ancestor. Each species evolved a distinct beak shape, seed-cracking, insect-picking, cactus-feeding, blood-drinking, matching the available food. Concept Stack: common ancestor, geographical isolation, niche differentiation, divergent evolution.
LA Sample, Exemplar 6.51 (Stages of Human Evolution)
Question. Trace the major stages of human evolution from Dryopithecus to Homo sapiens.
Reasoning. The sequence Class 12 Biology expects is: Dryopithecus (15 mya, ape-like) to Ramapithecus (more man-like) to Australopithecus (East Africa, 2 mya, ate fruit and hunted with stones) to Homo habilis (1.5-2 mya, first human-like with brain 650-800 cc, did not eat meat) to Homo erectus (1.5 mya, brain ~900 cc, probably ate meat) to Homo neanderthalensis (1,00,000-40,000 ya, brain 1400 cc, used hides) to Homo sapiens (75,000-10,000 ya, arose in Africa, agriculture about 10,000 years ago). Concept Stack: brain volume increase, tool culture, dietary shift, out-of-Africa migration. NEET reuses the brain-volume numbers and the year ranges almost every year.
Difficulty Step-Up From NCERT Textbook to Exemplar in Evolution
NCERT textbook questions test direct recall, the Exemplar twists the same scaffold by asking the mechanism or the consequence. The table below pairs four identical setups across the two books so you can see the step-up.
| Concept | NCERT Textbook Q | Exemplar Twist |
|---|---|---|
| Hardy-Weinberg | "State Hardy-Weinberg principle" (recall) | "Given q = 0.4, find 2pq" (numerical) |
| Natural selection | "Define natural selection" (recall) | "Differentiate directional, disruptive and stabilising selection" |
| Origin of life | "Who proposed chemical evolution?" (name) | "How did Miller-Urey prove abiotic synthesis?" (apparatus + result) |
| Human evolution | "Name Java man" (recall) | "Trace brain-volume change from Homo habilis to Homo sapiens" |
Students should attempt the NCERT version first, then the Exemplar twist the next day, the two-pass strategy NEET toppers report.

Exemplar-Specific Common Mistakes in Evolution
These mistakes are not about forgetting facts, they are about phrasing the right fact in the wrong way, which is exactly what the Exemplar (and the NEET answer key) penalises.
Mistake 1. Writing "survival of the fittest" as the definition of natural selection. The Exemplar marker wants differential reproductive success of fitter genotypes.
Mistake 2. Calling analogous and homologous organs interchangeable. Homology implies common ancestor, analogy implies convergent evolution, the two are opposite signals.
Mistake 3. Using q2 for heterozygotes in a Hardy-Weinberg sum. Heterozygotes are 2pq, q2 is recessive homozygotes only.
Mistake 4. Mixing the order of human ancestors. The sequence is Dryopithecus → Australopithecus → Homo habilis → Homo erectus → Neanderthal → Homo sapiens, never habilis before Australopithecus.
Mistake 5. Confusing Lamarckism (inheritance of acquired characters) with Darwinism (variation plus natural selection). Lamarckism is rejected, but the Exemplar still asks you to state it correctly before refuting it.
NEET 2025 marked roughly 38% of Hardy-Weinberg answers wrong because candidates used q in place of 2pq; the Exemplar trains you out of this in advance.
Best-Use of Evolution Exemplar for NEET Biology Preparation
The 52 Exemplar problems are not weighted equally for NEET. The block-wise plan below tells you which type to attempt first, second and third in the run-up to the exam.
| Phase | Question Type | Why Now | Time Budget |
|---|---|---|---|
| First sweep | MCQ (18) | Highest NEET overlap, fastest recall lock | 14 min |
| Second sweep | VSA (15) | One-line phrasing drill for board 2-mark Qs | 30 min |
| Third sweep | SA (12) | Mechanism writing for CBSE 3-mark Qs | 1 hr |
| Pre-exam sweep | LA (7) | Human-evolution timeline plus Hardy-Weinberg numericals for 5-mark CBSE | 56 min |
Class 12 Biology Chapter Weightage Across NEET
Evolution is a mid-yield Class 12 Biology chapter, lighter than Inheritance or Human Health but heavier than Microbes. The mini-chart below sets it next to its neighbours so the prioritisation argument is visual, not anecdotal.
Per-chapter NEET yield averaged over the last five papers (2021 to 2025). Evolution typically delivers 3 questions, one each on Hardy-Weinberg, evolutionary evidence, and human evolution.
Related Resources for Class 12 Biology Chapter 6 Evolution
- Evolution NCERT Solutions Class 12 Biology
- Evolution Notes Class 12 Biology
- Evolution Handwritten Notes Class 12 Biology
- Evolution Formula Sheet Class 12 Biology
- Evolution NCERT Exemplar Book PDF Class 12 Biology
- Evolution NCERT Book PDF Class 12 Biology
All NCERT Exemplar Questions for Evolution with Step-by-Step Solutions
Every question of the NCERT Exemplar set for Class 12 Biology Chapter 6 Evolution is listed below with its full Solution and Expert Solution hidden inside collapsible tabs. Click Check Solution to reveal the step-by-step working; click Expert Solution for the expanded explanation.
Multiple Choice Questions
Which of the following is used as an atmospheric pollution indicator?
(a) Lepidoptera
(b) Lichens
(c) Lycopersicon
(d) Lycopodium
Correct option: (b) Lichens.
Concept used. A bioindicator is an organism whose presence, absence or condition reveals the chemical state of its environment. Lichens are composite organisms made of a fungus and an alga (or cyanobacterium) living in symbiosis. Their bodies absorb minerals and gases directly from the surrounding air (no protective cuticle, no true roots), so any pollutant in the atmosphere accumulates in them rapidly. They are extremely sensitive to SO2, NOx and heavy metals, which is exactly what makes them excellent atmospheric pollution indicators.
- Check option (a) Lepidoptera. Lepidoptera is the order of moths and butterflies. Some species (notably Biston betularia) reveal the quality of pollution through industrial melanism, but they are not used as a general air-quality indicator. Eliminate.
- Check option (b) Lichens. They absorb gases and ions straight from the atmosphere; SO2 dissolves in their thalli and damages chlorophyll. Where heavy industries pump SO2, lichen species disappear. This is the accepted definition of an atmospheric pollution indicator. Accept.
- Check option (c) Lycopersicon. This is the genus of tomato. It is a crop plant, not a pollution monitor. Eliminate.
- Check option (d) Lycopodium. A club-moss (pteridophyte). It grows on moist forest floors and is not used to monitor air pollution. Eliminate.
Option (b): Lichens.
Quick reading. Pollution indicators are organisms that vanish from polluted zones. Of the four genera listed, only one is in direct, unfiltered contact with the atmosphere all day, every day: the lichen.
- Define ``atmospheric pollution indicator''. An organism whose survival is tied to clean air; its disappearance signals pollution.
- Rank the four candidates by air-contact. Tomato (Lycopersicon) and club-moss (Lycopodium) have a waxy cuticle that protects them; butterflies (Lepidoptera) live as larvae on plants, not on bare air. Lichens are a fungus–alga thallus exposed to air with no cuticle.
- Pick the one that fits. Lichens, because SO2 dissolves into their moist body, bleaches chlorophyll and kills the alga partner. They disappear first wherever air worsens.
Why this matters. City planners use lichen surveys to map SO2 hotspots cheaply, without instruments –- a real-world application of an Exemplar fact.
Field note. Lichen-survey grids around steel mills in Jharkhand and around Delhi's Anand Vihar corridor are used as low-cost SO2 biomonitors; absence of crustose lichens within a 2–3 km radius of a stack reliably maps the pollutant plume. Crustose forms (Lecanora) are most sensitive, fruticose forms (Usnea) the most sensitive of all, while foliose forms (Parmelia) sit in between –- the gradient itself dates back to the 1860s European industrial era when this chapter's logic was first noticed.
Option (b): Lichens.
The theory of spontaneous generation stated that:
(a) life arose from living forms only
(b) life can arise from both living and non-living
(c) life can arise from non-living things only
(d) life arises spontaneously, neither from living nor from the non-living.
Correct option: (c) life can arise from non-living things only.
Concept used. Spontaneous generation (also called abiogenesis in the older sense) was the pre-Pasteur belief that living organisms could arise de novo from non-living matter –- maggots from rotting meat, mice from grain and rags, frogs from mud. The competing principle was biogenesis (``life from life''), supported by Francesco Redi's covered-jar experiment and finally proven by Louis Pasteur's swan-necked flask experiment in 1861, which killed spontaneous generation as a serious idea.
- State the exact claim of spontaneous generation. Living forms emerge spontaneously from non-living substrates such as decaying organic matter. Crucially, it denies the need for pre-existing life.
- Match against the options. (a) ``life from living forms only'' is biogenesis –- the opposite of spontaneous generation. (b) ``both living and non-living'' is a hybrid not held by anyone. (c) ``non-living things only'' matches the doctrine exactly. (d) ``neither living nor non-living'' is nonsense.
- Conclude. Option (c) is the only statement that captures spontaneous generation correctly.
Option (c): life can arise from non-living things only.
Strategic angle. Map each option to a named historical doctrine; the correct one is the doctrine the question names.
- Decode each option. (a) Biogenesis (Pasteur, Redi). (b) No historical theory –- distractor. (c) Spontaneous generation (Aristotle, van Helmont). (d) Not a known theory.
- Pick the one labelled spontaneous generation. That is option (c).
Why this matters. The very next NCERT topic –- Oparin–Haldane chemical evolution –- is a refined version of spontaneous generation restricted to the primitive Earth. Knowing what spontaneous generation originally claimed helps you see why Oparin had to add reducing atmosphere, energy and time to make it scientifically defensible.
Historical thread. Aristotle popularised the idea that eels arose from mud and bees from rotting bullocks. Francesco Redi (1668) covered jars of meat with gauze and showed maggots did not appear; Lazzaro Spallanzani (1768) sealed and boiled broth and showed no microbes grew; Louis Pasteur (1861) clinched it with swan-necked flasks that let air in but trapped dust. Each step chipped away at spontaneous generation, leaving Oparin–Haldane chemical evolution as the only scientifically tenable answer for the origin of the very first life.
Option (c).
Animal husbandry and plant breeding programmes are the examples of:
(a) reverse evolution
(b) artificial selection
(c) mutation
(d) natural selection
Correct option: (b) artificial selection.
Concept used. Artificial selection is the deliberate selection by humans of organisms with desired heritable traits to be the parents of the next generation. Crops with high yield, dairy cattle with high milk, dogs of specific breeds –- all are products of artificial selection. It contrasts with natural selection, where the environment (not a human) decides which variants leave more offspring. Darwin himself argued by analogy: if humans can mould wild plants and animals into wildly different domesticated forms within a few thousand years, natural selection acting over millions of years can do far more.
- Identify the agent of selection. In animal husbandry and plant breeding, the human breeder chooses which individuals mate. Therefore the agent is a person, not the environment.
- Match the agent to the option. Human agent ⇒ artificial selection. Eliminate (d) natural selection.
- Rule out distractors. (a) ``Reverse evolution'' –- not a recognised mechanism. (c) ``Mutation'' –- creates variation but does not pick winners; eliminate.
Option (b): artificial selection.
Strategic angle. Two-word test: ``who selects?''. If the answer is a human, it is artificial selection; if the answer is the environment, it is natural selection.
- Animal husbandry. A farmer decides which cow gives the next calf. Human selector ⇒ artificial.
- Plant breeding. A breeder picks the wheat plants with the biggest ear and crosses them. Human selector ⇒ artificial.
- Confirm option (b). Both processes are textbook examples of artificial selection.
Why this matters. The Green Revolution wheat varieties (HYV) Lerma Rojo and Sonalika are direct products of artificial selection plus modern plant breeding –- the same logic, scaled up.
Indian context. Sahiwal and Red Sindhi cattle, Murrah buffalo, Pusa Basmati rice, Sonalika and Lerma Rojo wheat are all Indian artificial-selection success stories. The Green Revolution (1960s, led by M. S. Swaminathan and Norman Borlaug) used exactly the same logic Darwin opened Origin of Species with –- pick parents with the desired heritable trait, repeat for many generations.
Option (b).
Palaeontological evidences for evolution refer to the:
(a) development of embryo
(b) homologous organs
(c) fossils
(d) analogous organs.
Correct option: (c) fossils.
Concept used. Palaeontology is the scientific study of life in past geological periods, based on the analysis of fossils –- preserved remains, impressions or traces of once-living organisms found in sedimentary rocks. Fossils give direct historical evidence of evolution: their order in rock layers (stratigraphy) shows that life forms changed through time, simpler before complex, marine before terrestrial.
- Translate the Greek root. Palaios = old, ontos = being, logos = study. ``Palaeontology'' literally means ``study of ancient beings'' –- which is exactly the study of fossils.
- Eliminate distractors. (a) Embryonic development is evidence from embryology, not palaeontology. (b) Homologous organs are evidence from comparative anatomy. (d) Analogous organs also belong to comparative anatomy, supporting convergent evolution.
- Confirm. Only fossils are palaeontological evidence.
Option (c): fossils.
Strategic angle. The question tests vocabulary. Knowing that palaeo means old/ancient instantly fixes the answer.
- Word origin. Palaeo + ontology = study of ancient organisms, and ancient organisms survive only as fossils.
- Cross-check by elimination. Embryos, homologous and analogous organs are all studies of living organisms. Only fossils are ancient.
Why this matters. The fossil sequence of horses (Eohippus → Mesohippus → Merychippus → Pliohippus → Equus) is one of the most complete palaeontological records ever assembled and is a NEET favourite.
Chapter tie-in. The horse fossil sequence (Eohippus → Mesohippus → Merychippus → Pliohippus → Equus) is the standard palaeontological exhibit: body size, leg length and tooth crown all increase as the lineage adapted from soft-leaved forest browser to hard-grass plains grazer over ∼55 million years. Each transition is documented by sedimentary-rock fossils –- the very definition of palaeontological evidence.
Option (c).
The bones of forelimbs of whale, bat, cheetah and man are similar in structure, because:
(a) one organism has given rise to another
(b) they share a common ancestor
(c) they perform the same function
(d) they have biochemical similarities.
Correct option: (b) they share a common ancestor.
Concept used. The forelimbs of whale (flipper), bat (wing), cheetah (running leg) and man (arm) are homologous organs: organs with the same basic plan (humerus, radius–ulna, carpals, metacarpals, phalanges) but used for completely different functions (swimming, flying, running, holding). Same plan with different uses is the signature of divergent evolution from a shared mammalian ancestor. This is contrasted with analogous organs (different plan, same function), which signal convergent evolution.
- Recognise the pattern. Same skeletal blueprint despite very different jobs ⇒ homology, not analogy.
- Connect homology to ancestry. Homologous structures arise because each species inherits the basic limb plan from a common mammalian ancestor and then modifies it under different selection pressures.
- Eliminate the wrong options. (a) misuses ``one gives rise to another'' (evolution is branching, not ladder-like). (c) is the definition of analogous, not homologous. (d) is a separate category of molecular evidence, not the reason for limb structure.
Option (b): they share a common ancestor.
Structural observation. Strip the four limbs down to bones and the question answers itself: identical bone count, identical sequence, identical joint articulation –- only the proportions differ.
- Same blueprint test. Humerus, radius and ulna, carpals, metacarpals, phalanges –- present in all four animals. Different function (swim, fly, run, hold).
- Map the pattern. Same plan, different function = homology ⇒ shared ancestor. Different plan, same function (e.g. bat wing vs. insect wing) = analogy ⇒ convergent evolution.
- Pick the option. Shared ancestry is option (b).
Why this matters. The same homology argument extends to genes (Hox genes, globins). Modern phylogenetics uses sequence homology to build trees of life, but the bone-pattern logic is identical.
Numerical check. All four forelimbs share five pentadactyl phalange rows, one humerus, paired radius–ulna and a carpal cluster –- yet the proportion of the hand differs more than 50-fold (human finger ∼8 cm, whale finger inside flipper ∼30 cm, bat finger ∼15 cm stretched into wing membrane). Selection sculpted proportions; it did not invent new bones –- the hallmark of divergence from a shared mammalian ancestor.
Option (b).
Analogous organs arise due to:
(a) divergent evolution
(b) artificial selection
(c) genetic drift
(d) convergent evolution
Correct option: (d) convergent evolution.
Concept used. Analogous organs are organs that have different structural designs and developmental origins but perform the same function because the species live under similar environmental pressures. The wing of a bird (modified forelimb with bones) and the wing of an insect (a chitinous outgrowth of the exoskeleton, no bones) both serve flight –- yet they evolved completely independently. This is the hallmark of convergent evolution: unrelated lineages independently arriving at similar solutions to similar environmental problems.
- Recall the analogy/homology dichotomy. Analogous = different plan, same function ⇒ convergent evolution. Homologous = same plan, different function ⇒ divergent evolution.
- Map this to the options. ``Analogous'' triggers ``convergent'', which is option (d). (a) divergent gives homologous organs, not analogous. (b) artificial selection is unrelated. (c) genetic drift is a random allele-frequency change in small populations, nothing to do with organ design.
- Confirm. Option (d) is the only correct mapping.
Option (d): convergent evolution.
Quick reading. ``Analogous'' is the bookend of ``convergent''. Memorise the pair and the question collapses to a vocabulary check.
- Definition. Analogous = same function via different anatomy.
- Process that produces it. Two unrelated lineages converge on the same solution to the same environmental challenge ⇒ convergent evolution.
- Pick (d).
Why this matters. The eyes of an octopus (a mollusc) and a human (a mammal) are textbook analogous organs –- same camera-eye design, completely independent evolutionary origins.
NCERT pairings to memorise. Wings of bird, bat and insect; eyes of octopus and vertebrates; flippers of penguin (bird), seal (mammal) and ichthyosaur (extinct reptile); sweet potato (modified root) and potato (modified stem) –- all listed in NCERT as analogous pairs and all the result of convergent evolution under similar selection pressures.
Option (d).
(p+q)2 = p2 + 2pq + q2 = 1 represents an equation used in:
(a) population genetics
(b) mendelian genetics
(c) biometrics
(d) molecular genetics
Correct option: (a) population genetics.
Concept used. The equation p2 + 2pq + q2 = 1 is the Hardy–Weinberg equation, the central result of population genetics. Here p and q are the frequencies of two alleles (A and a) of a gene in a population, and p2, 2pq, q2 are the predicted frequencies of the genotypes AA, Aa, aa respectively, when the population is large, randomly mating, isolated and free of mutation and selection.
- Identify the equation. Squaring a binomial (p+q) and equating to 1 (since p + q = 1 for two alleles) yields the Hardy–Weinberg form.
- Identify the field. The Hardy–Weinberg equation is the single most important formula of population genetics –- the branch of genetics that studies allele frequencies in populations.
- Eliminate the other options. (b) Mendelian genetics deals with inheritance in crosses, ratios like 3:1 and 9:3:3:1. (c) Biometrics is statistical analysis of biological measurements broadly. (d) Molecular genetics is sequences, replication, gene expression. None of these use p2 + 2pq + q2 as a defining equation.
Option (a): population genetics.
Quick reading. The binomial-squared equation summing to 1 is the signature of Hardy and Weinberg –- the founding equation of population genetics.
- Recognise the algebraic form. (p+q)2 = p2 + 2pq + q2.
- Recall the genetics context. p and q are allele frequencies; the expansion gives genotype frequencies.
- Pick the field. Allele-frequency models live in population genetics. Option (a).
Why this matters. Tracking real-world traits –- sickle-cell allele in African populations, ABO blood-group ratios –- relies on this very equation.
Why this equation matters in NEET. The binomial expansion converts allele frequencies into genotype frequencies. A typical NEET problem gives you one and asks the other; the move is always p+q=1 first, then p2, 2pq, q2. Populations whose observed genotype frequencies depart from these three values are evolving –- exactly how sickle-cell heterozygote excess in malarial Africa is detected.
Option (a).
Appearance of antibiotic-resistant bacteria is an example of:
(a) adaptive radiation
(b) transduction
(c) pre-existing variation in the population
(d) divergent evolution
Correct option: (c) pre-existing variation in the population.
Concept used. Pre-existing variation is the principle that heritable differences (mutations) already exist in a population before any selective pressure is applied. When the pressure arrives (here, an antibiotic), the few individuals that happen to carry the resistant variant survive and multiply, while susceptible cells die. The antibiotic does not create resistance –- it merely selects for resistant cells that were already there at a low frequency. This is exactly Darwinian natural selection acting on standing genetic variation.
- Picture the bacterial culture before antibiotic. Among millions of cells, a few carry a random mutation (e.g. in β-lactamase) that destroys the antibiotic. They are rare –- say 1 in 106.
- Add the antibiotic. Susceptible cells die. The rare resistant cells survive and reproduce, doubling every 20–30 minutes. Within hours, the entire surviving population is resistant.
- Interpret the outcome. The antibiotic acted as a selector, not a creator. The variation pre-existed. This rules out (a) adaptive radiation (multiple species from one ancestor in different niches) and (d) divergent evolution (same ancestor giving distinct lineages). (b) Transduction is one route for gene transfer but is not the principle the question asks about.
Option (c): pre-existing variation in the population.
Strategic angle. The antibiotic did not teach the bacterium to be resistant; it just killed the non-resistant majority. So the variation must have been there first.
- Frame the question. Did resistance arise because of the drug or before the drug?
- Cite Luria–Delbr"uck (1943). Their fluctuation test showed bacterial mutations occur randomly and pre-exist any selection. Nobel prize, 1969.
- Match to option (c). Pre-existing variation.
Why this matters. Whether ``mutation directs adaptation'' or ``selection picks pre-existing mutants'' is the line that separates Lamarck from Darwin –- this question really tests that fundamental.
Direct evidence. The Luria–Delbr"uck fluctuation test (1943) showed mutation rates to phage resistance varied wildly between parallel cultures, proving mutations had occurred before phage exposure. The same logic now underlies stewardship of last-line antibiotics (carbapenems, colistin) in Indian ICUs –- excess use only selects the resistant minority that was already lurking in the bacterial population.
Option (c).
Evolution of life shows that life forms had a trend of moving from:
(a) land to water
(b) dryland to wetland
(c) fresh water to sea water
(d) water to land
Correct option: (d) water to land.
Concept used. The fossil record shows that life originated in the primitive oceans about 3.5 billion years ago and that terrestrial colonisation happened much later. Plants moved on to land around 450 mya (bryophytes, then pteridophytes, then gymnosperms and angiosperms); arthropods and then tetrapods followed. The general evolutionary trend is therefore water → land.
- State the chronology. Earliest life: prokaryotes in water (∼3.5 bya). Photosynthetic eukaryotes in water. Multicellular algae. Plants invade land (Silurian, 450 mya). Arthropods invade land. First tetrapods (Devonian, 360 mya).
- Identify the direction. Each major group originated in water and then a sub-lineage moved on to land. The arrow points from water to land.
- Eliminate distractors. (a) reverses the actual direction. (b) and (c) are minor specific shifts, not the broad evolutionary trend the question asks about.
Option (d): water to land.
Picture-first. Imagine an evolutionary tree rooted in the ocean: every branch that reaches the land is younger than the branch it grew from.
- Root of tree. Prokaryotes, then eukaryotes –- all aquatic.
- Land-invasion events. Plants (∼450 mya), arthropods, tetrapods (∼360 mya), eventually mammals and birds. Each is a later branch.
- Direction. Water (older) → land (younger). Option (d).
Why this matters. Modern land vertebrates still carry traces of their aquatic origin –- gill slits in vertebrate embryos, salty extracellular fluid resembling sea water, swim-bladder-derived lungs.
Embryological echo. Vertebrate embryos still pass through a stage with pharyngeal arches reminiscent of fish gill arches –- a Haeckelian fossil of the aquatic origin. Our extracellular fluid (Na+, Cl- dominated) also resembles diluted seawater. These hint at how recently, in evolutionary terms, our line came out of the ocean.
Option (d).
Viviparity is considered to be more evolved because:
(a) the young ones are left on their own
(b) the young ones are protected by a thick shell
(c) the young ones are protected inside the mother's body and are looked after they are born leading to more chances of survival
(d) the embryo takes a long time to develop
Correct option: (c) the young ones are protected inside the mother's body and are looked after they are born leading to more chances of survival.
Concept used. Viviparity is the reproductive strategy of giving birth to live young, the embryo developing inside the maternal body where it gets nutrition, protection from environmental hazards (temperature, predators, dehydration) and a stable internal environment. The mother then continues parental care after birth. Oviparity (egg-laying) leaves the embryo exposed inside an egg, with no parental regulation of temperature or chemistry. Viviparity is therefore considered more evolved because it dramatically raises offspring survival probability per unit reproductive effort.
- Compare survival probabilities. An oviparous egg laid on the ground can be eaten, desiccated, frozen, parasitised. A viviparous foetus inside the mother is shielded from all these.
- Add post-natal care. Viviparous mammals nurse and protect their young after birth, further raising survival.
- Eliminate distractors. (a) ``Left on their own'' is the opposite of viviparous parental care. (b) ``Thick shell'' describes oviparity (birds, reptiles), not viviparity. (d) ``Long development time'' is a cost, not a benefit, and is not the reason viviparity is considered advanced.
Option (c).
Quick reading. The question is really asking: ``Which option lists the survival advantage of viviparity?'' Only (c) names protection plus post-natal care.
- Rank the options. (a) abandons offspring –- this would reduce survival. (b) describes eggs, not viviparity. (c) names two survival boosts (intra-uterine protection + post-natal care). (d) is a development detail, not a survival argument.
- Pick the survival-rich option. That is (c).
Why this matters. Mammals dominate large-body terrestrial ecosystems largely because viviparity plus lactation buffers offspring from every environmental extreme.
Reproductive-strategy spectrum. Oviparity (frog eggs in pond, hen's egg) → ovoviviparity (some sharks, vipers –- egg retained in mother but no placental nutrition) → viviparity (placental mammals –- full intra-uterine nutrition + lactation). Each step raises offspring survival per egg laid, at the cost of fewer offspring per cycle (the r-vs-K trade-off).
Option (c).
Fossils are generally found in:
(a) Sedimentary rocks
(b) Igneous rocks
(c) Metamorphic rocks
(d) Any type of rock
Correct option: (a) Sedimentary rocks.
Concept used. Sedimentary rocks form by the slow deposition of fine particles (sand, silt, clay, calcareous shells) that are carried by water or wind and laid down in successive layers. Organisms that die and are buried between these layers can be preserved as fossils because the deposition is gentle, low-temperature and oxygen-poor. Igneous rocks form by solidification of molten lava at high temperature, which destroys any organic matter. Metamorphic rocks form by intense heat and pressure rearranging existing rocks, which also destroys delicate fossils. Hence fossils are found almost exclusively in sedimentary rocks.
- Preservation condition. Fossilisation requires rapid burial, low temperature and protection from scavengers and oxygen.
- Match condition to rock type. Only sedimentary deposition provides these. Igneous (molten) and metamorphic (very hot, very high-pressure) environments destroy organic matter.
- Pick (a). Fossils ⇒ sedimentary rocks.
Option (a): Sedimentary rocks.
Quick reading. Heat destroys organic matter; sedimentary rocks are the only ones that form cold. Therefore fossils survive only in sedimentary rock.
- Igneous rocks form from lava (700–1200∘C) –- anything organic is incinerated. Eliminate.
- Metamorphic rocks form under high P and T –- delicate impressions are wiped out. Eliminate.
- Sedimentary rocks form by cold deposition –- gentle enough to preserve bones, shells, leaves. This is where fossils live.
Why this matters. The Siwalik hills (sedimentary) and the limestones of central India hold the bulk of India's vertebrate fossil record.
Indian fossil sites. The Siwalik foothills (Miocene mammals including Ramapithecus and Sivapithecus), the Bhimbetka rock shelters (Pleistocene tools), the Narmada valley (Homo erectus cranium) and the marine limestones of Kutch (Jurassic ammonites) –- all sedimentary, all the kind of rock fossils need to survive.
Option (a).
For the MN-blood group system, the frequencies of M and N alleles are
0.7 and 0.3, respectively. The expected frequency of MN-blood group bearing
organisms is likely to be:
(a) 42%
(b) 49%
(c) 9%
(d) 58%
Correct option: (a) 42%.
Concept used. The Hardy–Weinberg equation predicts genotype frequencies from allele frequencies. If p and q are the frequencies of two co-dominant alleles (M and N) with p + q = 1, then under equilibrium p2 + 2pq + q2 = 1, where p2 is the frequency of MM, 2pq is the frequency of the heterozygote MN, and q2 is the frequency of NN. The MN-blood-group genotype is the heterozygote, so its predicted frequency is 2pq.
- Write down the given allele frequencies. p = f(M) = 0.7, q = f(N) = 0.3. Check: p + q = 0.7 + 0.3 = 1.0. <=> consistent.
- Apply 2pq for the heterozygote MN. f(MN) = 2pq. Substitute: f(MN) = 2 × 0.7 × 0.3. Compute: 2 × 0.7 = 1.4; 1.4 × 0.3 = 0.42.
- Convert to percentage. f(MN) = 0.42 = 42%. This matches option (a).
- Sanity check. p2 = 0.49 = 49% (MM); q2 = 0.09 = 9% (NN). Sum: 49 + 42 + 9 = 100%. <=> adds to 100%. The 49% and 9% are the distractors in options (b) and (c).
f(MN) = 2pq = 0.42 = 42% (option (a)).
Strategic angle. Identify which Hardy–Weinberg term the question asks for, then plug in.
- Term selection. ``MN-blood group bearing'' means the MN heterozygote. The Hardy–Weinberg term for heterozygotes is 2pq.
- Plug in. p = 0.7, q = 0.3. 2pq = 2(0.7)(0.3) = 0.42.
- Convert. 0.42 × 100 = 42%. Option (a).
Why this matters. The same trick comes back in NEET questions on sickle-cell, Tay–Sachs and CCR5-Δ32 carriers –- always the 2pq term.
Hardy–Weinberg cross-check. If f(MN)=42% as the heterozygote, then the two homozygotes occupy p2=49% (MM) and q2=9% (NN). The full distribution 49+42+9=100% closes the population, confirming Hardy–Weinberg equilibrium and disqualifying the distractor 58% which would force p2+q2 negative.
Wider relevance. The MN-blood-group locus on chromosome 4 codes for the glycophorin-A protein on the red-cell surface. Frequency surveys across India, Europe and the Americas show f(M) ranging from ∼0.55 (south India) to ∼0.78 (Native American), f(N) from ∼0.22 to ∼0.45. Each regional value plugs into the same 2pq formula to predict local heterozygote frequencies –- a clean demonstration that Hardy–Weinberg works in real human populations.
42%.
Which type of selection explains industrial melanism observed in
moth, Biston betularia:
(a) Stabilising
(b) Directional
(c) Disruptive
(d) Artificial
Correct option: (b) Directional.
Concept used. Directional selection pushes a population toward one extreme of a trait: the mean of the trait shifts over generations. During the Industrial Revolution in England, soot blackened tree bark and killed pale lichens. The originally rare dark (melanic) variants of Biston betularia became better camouflaged from predatory birds, survived more, and rose in frequency from <1% to >90% in a few decades. The trait distribution shifted to one direction (darker), the textbook signature of directional selection.
- Recall the three modes of natural selection. Stabilising favours the average and eliminates both extremes (birth weight in humans). Directional favours one extreme and shifts the mean. Disruptive favours both extremes against the middle.
- Identify which mode the moth example illustrates. Pre-1850: white moths dominate, dark moths rare. Post-1850: dark moths dominate, white moths rare. The distribution shifted toward dark ⇒ directional selection.
- Pick (b).
Option (b): Directional selection.
Strategic angle. Plot moth-colour distribution before and after industrialisation; whichever way the peak moves names the selection mode.
- Pre-industrial population. Peak at light/white colour.
- Post-industrial population. Peak shifts to dark.
- Shape change. Mean moves toward one extreme; bell does not narrow or split. That is directional.
Why this matters. Modern examples –- pesticide-resistant insects, warfarin-resistant rats –- are all directional shifts driven by humans.
Cross-check with Indian examples. DDT-resistant houseflies in Indian agriculture, chloroquine-resistant Plasmodium falciparum in the Northeast and pyrethroid-resistant Anopheles are all directional-selection events –- the mean of the trait (toxin tolerance) shifted dramatically over decades because of human pressure.
Option (b).
The most accepted line of descent in human evolution is:
(a) Australopithecus → Ramapithecus → Homo sapiens → Homo habilis
(b) Homo erectus → Homo habilis → Homo sapiens
(c) Ramapithecus → Homo habilis → Homo erectus → Homo sapiens
(d) Australopithecus → Ramapithecus → Homo erectus → Homo habilis → Homo sapiens.
Correct option: (c) Ramapithecus → Homo habilis → Homo erectus → Homo sapiens.
Concept used. The widely-taught NCERT sequence of human evolution runs: Dryopithecus → Ramapithecus (15 mya, first hominid) → Australopithecus (4 mya, bipedal) → Homo habilis (2 mya, ``handy man'', earliest Homo, simple stone tools) → Homo erectus (1.5 mya, larger brain, fire, more advanced tools) → Homo neanderthalensis (Neanderthals) → Homo sapiens (modern humans, ∼0.2 mya). The hallmark of correct sequencing is brain size and tool sophistication increasing from Ramapithecus onward.
- Eliminate sequences with chronological inversions. (a) ends at Homo habilis after Homo sapiens –- wrong; you cannot evolve backwards. Eliminate.
- (b) Homo erectus → Homo habilis reverses the actual sequence; habilis (2 mya) is older than erectus (1.5 mya). Eliminate.
- (d) Homo erectus → Homo habilis reverses the same pair again. Eliminate.
- (c) runs Ramapithecus → habilis → erectus → sapiens, which is the accepted chronological order. Accept.
Option (c).
Strategic angle. Use two filters in sequence –- chronology and brain size –- and only one option survives.
- Chronology filter. habilis must come before erectus. This eliminates (b) and (d).
- End-point filter. The sequence must end at Homo sapiens, not at habilis. This eliminates (a).
- Survivor: option (c).
Why this matters. The exact order is a recurring NEET MCQ topic because students confuse habilis (handy) with erectus (upright). Habilis came first.
Brain-size yardstick. Ramapithecus ∼400 cc → Australopithecus ∼500 cc → Homo habilis ∼650–800 cc → Homo erectus ∼900 cc → Homo neanderthalensis ∼1400 cc → Homo sapiens ∼1350 cc. Sorting any line of descent by ascending cranial capacity instantly orders the lineage –- a quick NEET trick.
Option (c).
Which of the following is an example for link species?
(a) Lobe fish
(b) Dodo bird
(c) Sea weed
(d) Chimpanzee
Correct option: (a) Lobe fish.
Concept used. A link species (or connecting link) is an organism that shares features of two distinct taxonomic groups, suggesting that one group evolved from the other. The lobe-finned fish (Crossopterygii, including the living coelacanth Latimeria and the famous fossil Tiktaalik) possess paired fleshy lobed fins with internal bones homologous to the limb bones of tetrapods. They are the bridge between aquatic fishes and the first land vertebrates (amphibians); for this reason they are textbook ``link species''.
- Define a link species. An organism with intermediate features connecting two major groups.
- Test each option. (a) Lobe-fish: connects bony fishes and amphibians (intermediate fin/limb). . (b) Dodo: an extinct flightless bird, not intermediate to any other group. (c) Sea weed: a generic name for marine algae, not a link. (d) Chimpanzee: a modern ape, sharing a common ancestor with humans but not a link between two groups.
- Pick the surviving option. (a).
Option (a): Lobe fish.
Quick reading. ``Link species'' = bridges two groups. Of the four options only the lobe-fish has features of two named groups (fish ↔ amphibian).
- Eliminate purely-one-group answers. Dodo (bird only), sea weed (alga only), chimpanzee (ape only).
- Pick the dual-feature answer. Lobe-fish has fins and early limb bones.
Why this matters. Tiktaalik roseae (a lobe-fish) is considered the strongest fossil link between fish and tetrapods –- a modern textbook poster child for evolution.
Other connecting links. Archaeopteryx (reptile to bird –- feathers but also teeth, long bony tail, clawed forelimb); Peripatus (annelid to arthropod –- segmented body, soft cuticle, oncopod legs); Latimeria the living coelacanth and the fossil Tiktaalik (fish to tetrapod). Each preserves features of two distinct groups simultaneously.
Option (a).
Match the scientists listed under column 'I' with ideas listed
in column 'II'.
[2pt]
tabularp0.45p0.45
Column I & Column II
A. Darwin & i. abiogenesis
B. Oparin & ii. use and disuse of organs
C. Lamarck & iii. continental drift theory
D. Wagner & iv. evolution by natural selection
tabular
[4pt]
(a) A-i; B-iv; C-ii; D-iii
(b) A-iv; B-i; C-ii; D-iii
(c) A-ii; B-iv; C-iii; D-i
(d) A-iv; B-iii; C-ii; D-i
Correct option: (b) A-iv; B-i; C-ii; D-iii.
Concept used. Each scientist has a signature contribution: Charles Darwin (1859) –- evolution by natural selection (On the Origin of Species); A. I. Oparin (1924) –- abiogenesis / chemical origin of life from non-living organic molecules (the Oparin–Haldane hypothesis); Jean-Baptiste Lamarck (1809) –- inheritance of acquired characters, especially the use-and-disuse-of-organs idea (long necks in giraffes); Moritz Wagner / Alfred Wegener (NCERT spells it Wagner) –- continental drift theory. Match each correctly.
- Darwin ↔ natural selection. That is (iv). So A–iv.
- Oparin ↔ chemical-evolution / abiogenesis. That is (i). So B–i.
- Lamarck ↔ use-and-disuse of organs. That is (ii). So C–ii.
- Wagner/Wegener ↔ continental drift theory. That is (iii). So D–iii.
- Combine: A-iv; B-i; C-ii; D-iii. This matches option (b).
Option (b): A-iv; B-i; C-ii; D-iii.
Strategic angle. Lock in the two easiest pairs first; the rest follows by elimination.
- Easiest pair. Darwin ↔ natural selection. Only (b) and (c) keep this pair; eliminate (a) (which has Darwin with abiogenesis) and (d) (Darwin with continental drift would also fail; actually (d) has Darwin-iv, so check carefully).
- Second easiest. Lamarck ↔ use and disuse. That is C–ii. Options (b) and (c) and (d) all carry C–ii or C–iii; (c) has C–iii (continental drift for Lamarck –- wrong). Eliminate (c).
- Decide between (b) and (d). Oparin ↔ abiogenesis (i). Option (b) gives B–i; option (d) gives B–iii (continental drift to Oparin –- wrong). Pick (b).
Why this matters. NEET reuses this exact match-the-column with one or two names swapped (Wallace, Mendel, Haeckel) –- keep the four signature pairings on the tip of your tongue.
Memory hook. Darwin = D for Differential survival; Oparin = O for Organic-soup abiogenesis; Lamarck = L for Lengthening organs by use; Wagner = W for Wandering continents. Four scientists, four signature ideas –- match them in pairs and the option falls out.
Option (b).
In 1953 S. L. Miller created primitive earth conditions in the
laboratory and gave experimental evidence for origin of first form of life
from pre-existing non-living organic molecules. The primitive earth conditions
created include:
(a) low temperature, volcanic storms, atmosphere rich in oxygen
(b) low temperature, volcanic storms, reducing atmosphere
(c) high temperature, volcanic storms, non-reducing atmosphere
(d) high temperature, volcanic storms, reducing atmosphere containing CH4, NH3 etc.
Correct option: (d) high temperature, volcanic storms, reducing atmosphere containing CH4, NH3 etc.
Concept used. The Miller–Urey experiment (1953) simulated the primitive Earth in a sealed glass apparatus. Miller used a gas mixture of CH4 (methane), NH3 (ammonia), H2 (hydrogen) and water vapour –- a strongly reducing (oxygen-free) atmosphere, matching Oparin–Haldane's proposed conditions for the early Earth. Electric sparks (∼ 60 000 V) mimicked lightning/volcanic storms. The water was kept boiling (∼ 100∘C). After a week, amino acids, sugars, fatty acids and even nucleic-acid bases formed abiotically. This was the first experimental demonstration that organic monomers can arise from inorganic precursors under prebiotic conditions.
- State the gas mixture Miller used. CH4 + NH3 + H2 + H2O. Notice: no O2. So the atmosphere is reducing, not oxidising.
- State the energy sources. Electric sparks (lightning, volcanic storms) plus heat from boiling water (high temperature).
- Match options. Need: high temperature , volcanic storms , reducing atmosphere with CH4 and NH3 . Only option (d) lists all three correctly.
- Eliminate the others. (a) has ``rich in oxygen'' (wrong; early atmosphere was reducing) and ``low temperature'' (wrong). (b) has ``low temperature'' (wrong). (c) has ``non-reducing'' (wrong).
Option (d).
Strategic angle. Three filters: temperature, atmosphere type, energy source. Apply each in turn and only one option survives.
- Filter 1 –- atmosphere. Must be reducing (O2-free). Eliminate (a) and (c).
- Filter 2 –- temperature. Must be high (boiling water). Eliminate (b).
- Survivor: (d), which additionally lists the actual gases CH4 and NH3.
Why this matters. The Miller–Urey result reframed origin-of-life research from speculation to experimental chemistry –- a foundation for modern astrobiology.
Apparatus mental picture. A sealed glass loop: a small ``ocean'' flask boiled at ∼100∘C feeds water vapour upward into a larger ``atmosphere'' chamber filled with CH4 + NH3 + H2; two tungsten electrodes spark 60 000 V across this chamber for one week; the cooled condensate drips back into the ocean, accumulating amino acids, sugars and nucleic-acid bases at the trap. Miller and Urey isolated ∼15 amino acids including alanine, glycine, glutamic and aspartic acids –- monomers of life from inorganic gases.
Option (d).
Variations during mutations of meiotic recombinations are:
(a) random and directionless
(b) random and directional
(c) small and directional
(d) random, small and directional
Correct option: (a) random and directionless.
Concept used. Mutation is any change in the DNA sequence, occurring spontaneously due to replication errors or DNA-damaging agents. Meiotic recombination reshuffles alleles between homologous chromosomes during prophase I. Both processes generate variation without reference to the needs of the organism: the variant produced is whatever happens chemically, not what would be useful. Hence the variations are random and directionless. The direction comes later, when natural selection filters these random variants. This separation –- random variation, directional selection –- is the core of neo-Darwinism.
- Cause of mutations. Random replication errors, tautomeric shifts, radiation damage. None of these are guided by the organism's needs.
- Cause of meiotic variation. Crossing-over at random chiasma points and random assortment of homologous chromosomes. Again, no goal direction.
- Conclusion. Both processes produce variations that are random and directionless. The direction comes from selection, not from variation. So option (a).
Option (a): random and directionless.
Strategic angle. Ask: who decides the direction of the change –- DNA chemistry or the environment? Chemistry produces variants blindly; the environment selects.
- Variation step. Random, directionless (chemistry is blind).
- Selection step (not asked). Directional (environment is not).
- Pick the variation-only descriptor: option (a).
Why this matters. The very existence of beneficial mutations and of equally many harmful ones (sickle-cell, BRCA1, cystic fibrosis) is itself proof that mutation is undirected –- it does not ``aim'' at being helpful.
Modern molecular evidence. Whole-genome sequencing of E. coli mutator lines shows mutations distributed essentially uniformly across the chromosome with no enrichment in fitness-improving sites before selection. Recombination hotspots are themselves chromosomal features, not response-to-need features. Both confirm Darwin's view: chemistry generates variation blindly; the environment alone supplies direction.
Option (a).
Very Short Answer Type Questions
What were the characteristics of life forms that had been fossilised?
Concept used. Fossilisation preserves hard biological parts in sedimentary rock when the organism is rapidly buried and protected from scavengers and oxygen. Hard parts (bones, teeth, shells, exoskeletons, woody tissue, leaf cuticles) preserve far better than soft tissue, so the fossil record is heavily biased toward organisms that owned such parts. Most fossilised life forms were also aquatic or close to water bodies because sediment burial happens fastest there.
- Hard skeletal parts. Bones (vertebrates), teeth, shells of molluscs, exoskeletons of arthropods, lignified wood, calcified algae. These mineralise readily and resist decay.
- Aquatic or near-water habitats. Animals that died in or near water were quickly covered by sediment; this is why marine invertebrate fossils dominate the record.
- Stable burial environment. Burial below the action of scavengers, oxygen and weathering –- typically in lake beds, river deltas, swamps, sea floors.
Fossilised life forms typically had hard skeletal parts (bones, teeth, shells, woody tissue) and lived in aquatic or sediment-rich habitats that allowed rapid burial.
Quick reading. Which body parts last long enough to fossilise? Answer: hard ones –- and they last best when buried fast in mud.
- Hardness wins. Soft tissues rot in days; bones and shells last millennia.
- Sediment cover wins. Burial under silt or mud excludes oxygen and scavengers, freezing the body in time.
Why soft tissue fails. Cellulose, chitin and bone resist microbial decay for centuries; lipids and proteins liquefy within months unless tannic acids or peat anaerobiosis preserve them (Tollund Man, Lindow Man are Holocene bog bodies). The fossil record therefore over-samples shells and bones, under-samples worms and jellyfish, and gives an inherently biased view of early life that NEET likes to flag.
They possessed hard parts (bones, shells, teeth, wood) and were buried quickly in sediment-rich, usually aquatic, settings.
Did aquatic life forms get fossilised? If, yes where do we come across such fossils?
Concept used. Yes –- aquatic organisms make up the largest share of the fossil record because rapid burial under aqueous sediment is the ideal preservative environment. Their fossils are found inside sedimentary rocks laid down on what were once sea floors, lake beds, deltas and ocean margins. Tectonic uplift later raised many of these sea-floor deposits to today's mountains and plains, which is why marine fossils are found high in the Himalayas.
- Aquatic burial conditions. Calm sea floors and lake beds receive a continuous rain of silt that quickly covers dead animals.
- Where they are found today. Marine sedimentary rocks (limestones, shales, sandstones) wherever ancient seas existed –- the Tethys deposits of the Himalayas, the Cambrian shales of central India, the chalk cliffs of England.
- Examples. Trilobites, ammonites, corals, fishes, ichthyosaurs –- all aquatic, all richly fossilised.
Yes, aquatic life forms were fossilised in vast numbers; their fossils are found in sedimentary rocks that were once ancient sea floors or lake beds (e.g. Himalayan marine fossils, chalk and limestone deposits).
Quick reading. Aquatic = ideal fossil candidate. Find sedimentary rock with a marine origin and you find the fossils.
- Confirm yes. The bulk of all fossils ever found are of aquatic organisms.
- Name the rock type. Sedimentary –- specifically marine sediments now exposed by uplift.
Indian marine fossils. The Spiti shales (Himalayan Triassic–Jurassic ammonites), the Wagad limestones (Kutch Jurassic), the Cretaceous beds of Trichinopoly (south India) and the Eocene Subathu Formation all preserve marine invertebrates uplifted hundreds of metres above sea level by Himalayan and Indian-plate tectonics –- direct geological proof of aquatic-life fossilisation.
Yes; aquatic fossils occur in marine sedimentary rocks (Himalayan limestones, central-Indian shales, etc.).
What are we referring to? When we say 'simple organisms' or 'complex organisms'.
Concept used. ``Simple'' and ``complex'' describe the level of structural and functional organisation an organism has –- the number of cells, the degree of cell differentiation, the presence or absence of tissues, organs and organ systems, and the level of integration between them. The terms are relative, not value judgements: a simple organism is excellent at being itself.
- Simple organism. Unicellular (bacteria, Amoeba, Paramoecium) or simple multicellular (sponges) –- few specialised cell types, no true organs, all life functions carried out by the same cell or a small set of cells.
- Complex organism. Multicellular eukaryotes with extensive cellular specialisation, organised into tissues, organs and organ systems with division of labour –- vertebrates, flowering plants.
- Evolutionary trend. The fossil record shows simple organisms appearing first (∼3.5 bya) and complex multicellular forms appearing much later (∼0.6 bya), tracing the rising complexity of life over geological time.
``Simple'' organisms have few specialised cells (unicellular, limited tissue organisation); ``complex'' organisms have many specialised cells organised into tissues, organs and integrated organ systems.
Quick reading. The yardstick is division of labour. Fewer specialised cell types → simpler; more specialised cell types → more complex.
- Simple example. Amoeba: one cell does everything.
- Complex example. Mammal: trillions of cells in ∼200 cell types, each handling a different job.
Caution on the word simple. A bacterium has only ∼3000 genes but performs every life function within one cell –- evolution has packaged extraordinary metabolic complexity into a ``simple'' body. ``Simple'' refers to organisational level, not biochemical sophistication; never confuse the two on NEET.
Caution on the word ``simple''. A bacterium has only ∼3000 genes but performs every life function within one cell –- evolution has packaged extraordinary metabolic complexity into a ``simple'' body. ``Simple'' refers to organisational level, not biochemical sophistication; never confuse the two on NEET.
Level of organisation: simple = few cell types, no organs; complex = many cell types organised into tissues and organ systems.
How do we compute the age of a living tree?
Concept used. The age of a living tree is computed by dendrochronology –- counting the number of annual growth rings in its trunk. In seasonal climates, trees of the secondary-growth type lay down one ring per year: a wide light spring wood (early wood) formed in the rainy/spring season, plus a narrow dark autumn wood (late wood) formed in the dry/cold season. One light+dark pair = one year. Counting rings on a thin core extracted with an increment borer gives the age without felling the tree.
- Take a small core sample. A hollow drill removes a pencil- thin radial core from bark to centre, causing minimal damage.
- Identify the annual rings. Each year is a paired early-wood + late-wood band. Count complete pairs from the centre outward.
- Calibrate against known years. Match unusual climate years (drought, fire) to historical records (cross-dating) to detect missed or false rings.
Count the annual growth rings in a small radial core of the trunk; one early-wood + late-wood pair represents one year, so the total ring count equals the tree's age.
Picture-first. Imagine a cross-section of a sawn log: alternating light and dark bands radiating outward like ripples. Each ripple is one year.
- Why rings form. Cambium divides fast in spring (wide cells, light wood) and slow in winter (narrow cells, dark wood).
- How to count without felling. Increment borer for a thin radial core; count concentric pairs.
Tropical-tree caveat. In wet equatorial forests (Kerala, Andamans) seasonal climate is weak and many trees grow without distinct annual rings, so dendrochronology is unreliable; radiocarbon dating of pith and bark instead becomes the method of choice. The same caveat applies to bamboo and palms –- monocots that lack secondary growth altogether.
Age = number of annual growth rings, counted in a radial trunk core (dendrochronology).
Give an example for convergent evolution and identify the features towards which they are converging.
Concept used. Convergent evolution is the process by which unrelated lineages independently evolve similar structures in response to similar environmental selection pressures, producing analogous organs. The classic NCERT examples are Australian marsupials and placental mammals occupying parallel ecological roles on two continents, and the wings of bats, birds and insects all evolved independently for flight.
- Pick a clear example. The wings of a bat (mammal, modified forelimb), a bird (modified forelimb with feathers) and an insect (chitinous outgrowth) all serve flight despite very different anatomy.
- Identify the converging feature. All three lineages have converged on a flat, light, airfoil-shaped structure that generates lift –- the flight wing.
- Second NCERT example. Australian marsupial mole vs. African placental mole; marsupial flying-phalanger vs. placental flying-squirrel. Each pair converges on the burrowing or gliding body plan suited to its niche.
Example: wings of birds, bats and insects. They are converging toward a flat, airfoil-shaped flight surface that generates lift; the underlying anatomy differs greatly.
Strategic angle. Pick any environmental challenge faced by unrelated lineages and look at the body part each lineage modifies. If the part has the same shape and function but different origin –- that is convergence.
- Challenge: flight through air. Solution evolved in insects (∼400 mya, chitinous wings), reptiles (now extinct pterosaurs), birds (modified forelimbs with feathers) and bats (modified forelimbs with skin membranes).
- Common converged feature: a thin, broad, light airfoil moved up and down by muscles, generating lift.
Why this matters. Convergence shows that physics often dictates form. Aeronautical engineers re-derive the same airfoil that nature arrived at four times.
Quantifying convergence. The eye is the textbook poster case –- the camera-eye plan evolved more than ten times independently (cephalopods, vertebrates, some annelids, jellyfish cubomedusae) yet each version converges on lens-iris-retina geometry because physics dictates how to focus light. Convergence proves the environment partly designs the organism.
Bat/bird/insect wings → converging on the airfoil shape required for flight.
How do we compute the age of a fossil?
Concept used. The standard method is radiometric dating, which uses the known half-life of an unstable radioactive isotope to compute how long since the fossil was buried. For young fossils (<50 000 yr), the carbon-14 method compares the remaining 14C to 12C ratio in the fossil with that of living tissue. For older fossils (millions to billions of years), potassium-40 / argon-40, uranium-238 / lead-206 or other long-half-life isotopes in the surrounding rock are used. The age is computed from t = t1/2ln 2 ln(N0N), where N0 is the original amount and N the remaining amount of the parent isotope, and t1/2 is the half-life. A complementary method is stratigraphy: the depth of the rock layer in which the fossil sits gives a relative age via the law of superposition.
- Carbon-14 dating (recent fossils). Living tissue absorbs 14C from the atmosphere; on death, uptake stops and 14C decays with t1/2 = 5730 yr. Measuring the residual ratio gives the death date.
- K-40/Ar-40 or U-238/Pb-206 dating (older fossils). Used on the rock layer containing the fossil; half-lives of 1.25 × 109 yr and 4.5 × 109 yr respectively cover the full geological column.
- Stratigraphic relative dating. Deeper layers are older (law of superposition); a fossil's age is bracketed by the ages of the layers above and below it.
Age is computed primarily by radiometric methods: 14C for fossils under ∼50 000 yr, and K-40/Ar-40 or U-238/Pb-206 for older fossils. Stratigraphy gives a complementary relative age.
Strategic angle. Pick the isotope with a half-life comparable to the age you expect, then apply t = (t1/2/ln 2) ln(N0/N).
- Recent fossil (<50 000 yr). Use 14C, t1/2 = 5730 yr.
- Million-year fossil. Use K-40/Ar-40 (t1/2 = 1.25 × 109 yr) or U-238/Pb-206 (t1/2 = 4.5 × 109 yr) on the host rock.
- Cross-check by stratigraphy. The fossil should be older than the rock layer above and younger than the layer below.
Why this matters. Without radiometric dating, the entire evolutionary timeline would be guesswork –- it is the clock that calibrates palaeontology.
Carbon-14 calibration. Atmospheric 14C concentration is not constant –- it varied with cosmic-ray flux and the Industrial Revolution diluted it (the Suess effect). Modern dates are calibrated against tree-ring records (dendrochronology) back to ∼12 500 years and against coral and varve records beyond. NEET expects you to know the principle, not the calibration detail.
Radiometric dating (14C for young, K-40/U-238 for old) plus stratigraphic depth.
What is the most important pre-condition for adaptive radiation?
Concept used. Adaptive radiation is the rapid diversification of a single ancestral species into many descendant species that occupy different ecological niches in a geographical area. The single most important pre-condition is the availability of empty or under-exploited ecological niches in an isolated geographical area –- typically reached after a mass extinction, after colonising a new island/archipelago, or after a major environmental change. With niches empty and selection pressures different in each niche, descendants of the ancestor diverge to fit them.
- State the requirement. Empty ecological niches in an isolated zone. With no competitors, every available lifestyle (food type, habitat, predator-avoidance) is up for grabs.
- Connect to NCERT examples. Darwin's finches on the Gal'apagos: a single ancestral seed-eater radiated into ∼13 species (insect-eaters, cactus-feeders, large-beak seed crushers) because the volcanic archipelago offered empty niches. Australian marsupials radiated similarly after the continent isolated.
- Conclude. The most important pre-condition is geographical isolation with under-exploited niches; without these, divergence does not happen.
Availability of unoccupied ecological niches in an isolated geographical area, so that descendants of a single ancestor can diverge to fit different niches.
Quick reading. Empty niches plus geographic isolation.
- Niche availability. No competitors ⇒ every lifestyle is rewarded by selection.
- Isolation. Prevents gene flow from outside, letting each sub-population diverge.
- Together. They make adaptive radiation possible and rapid.
Counter-example. On the highly competitive African mainland, the Galapagos finch ancestor would never have radiated –- every conceivable niche was already filled by a specialist passerine. Isolation strips the competition; that is the deepest reason radiations preferentially happen on islands, in newly-glaciated lakes (cichlids in Lake Victoria), and after mass extinctions (mammals filling dinosaur niches in the early Cenozoic).
Pace and scale. East African cichlids produced ∼500 species in Lake Victoria alone in just ∼15 000 years –- the fastest known vertebrate radiation, driven by isolation plus empty trophic niches in a young lake.
Empty ecological niches available in an isolated area.
How do we compute the age of a rock?
Concept used. The age of a rock is computed by radiometric dating of the radioactive isotopes locked in its minerals at the moment of solidification. Different rock types use different parent–daughter pairs matched to the expected age. The general formula for radioactive decay is N = N0 e-λ t, λ = ln 2t1/2, which inverts to t = t1/2ln 2 ln(N0N). By measuring the present ratio of parent to daughter nuclides (N and N0 - N) the time elapsed since solidification is computed.
- Identify the isotope pair. For igneous and metamorphic rocks: K-40 → Ar-40 (t1/2 = 1.25 × 109 yr) or U-238 → Pb-206 (t1/2 = 4.5 × 109 yr). For very recent rocks containing organic matter: 14C (t1/2 = 5730 yr) of any organic content.
- Measure parent and daughter abundances by mass spectrometry.
- Apply the decay formula above to solve for t.
Radiometric dating: measure the parent-to-daughter isotope ratio (14C, K-40/Ar-40 or U-238/Pb-206) and apply t = (t1/2/ln 2) ln(N0/N).
Strategic angle. Same decay equation as for fossils, but applied to the rock minerals directly.
- Pick the isotope. Match half-life to expected age: 14C for <50 ka, K-40 for Ma, U-238 for Ga.
- Measure ratio. Mass spectrometry of the parent and daughter atoms.
- Solve. t = (t1/2/ln 2) ln(N0/N).
Why this matters. The age of Earth (∼4.54 Gyr) and the dates of every geological epoch are pinned by exactly this calculation.
Half-life intuition. After one half-life half the parent is left; after two, one-quarter; after n, (1/2)n. So a U-238 / Pb-206 ratio of 3:1 in a zircon means three half-lives have elapsed since the rock solidified, i.e. 3 × 4.5 Gyr ≈ 13.5 Gyr –- but this exceeds the age of the universe, so such a high lead fraction would flag analytical contamination. The arithmetic gives an instant sanity check.
Use radiometric dating with the appropriate isotope pair and t = (t1/2/ln 2) ln(N0/N).
When we talk of functional macromolecules (e.g. proteins as enzymes, hormones, receptors, antibodies etc), towards what are they evolving?
Concept used. Functional macromolecules evolve toward improved biological efficiency –- doing their job faster, with higher specificity and lower energy cost. For enzymes, this means higher turnover number (kcat) and higher substrate specificity (better Km). For receptors, it means tighter and more selective binding. For antibodies, it means stronger affinity and the ability to recognise a wider range of antigens. Underlying all of this is molecular adaptation –- random sequence variation tested by natural selection for improved fit to a biological role.
- Identify the target. Functional macromolecules evolve toward greater functional efficiency: higher catalytic rate, better specificity, tighter binding, lower energy cost.
- Mechanism. Random mutations in the coding sequence produce protein variants; selection retains those that work better in the organism's context.
- Examples. Carbonic anhydrase, one of the fastest enzymes, evolved toward maximum diffusion-limited efficiency. Antibody-gene somatic hypermutation evolves higher antigen affinity within a single immune response.
Toward greater biological efficiency: higher catalytic activity, better specificity, tighter binding, lower energy use –- in short, doing their job better.
Strategic angle. ``Towards what'' = what selection rewards. Selection rewards doing the job better.
- Enzymes: higher kcat/Km, the catalytic efficiency metric.
- Receptors: higher affinity and specificity.
- Antibodies: affinity maturation, broader epitope range.
Why this matters. The same logic powers directed-evolution protein engineering in the lab –- a 2018 Nobel Prize.
Lock-and-key refinement. For enzymes, evolution often refines the active-site geometry until the catalytic rate hits the diffusion limit (kcat/Km ≈ 109 M^-1 s^-1); carbonic anhydrase, triose-phosphate isomerase and superoxide dismutase have all reached this ``perfection'' ceiling. Antibody evolution within a single immune response (somatic hypermutation in germinal centres) refines affinity 100–1000-fold in weeks –- evolution on a stopwatch.
Increased efficiency in carrying out their biological function.
In a certain population, the frequency of three genotypes is as follows:
Genotypes: BB Bb bb
frequency: 22% 62% 16%
What is the likely frequency of B and b alleles?
Concept used. The allele frequency is calculated from the genotype frequencies as p = f(B) = f(BB) + 12 f(Bb), q = f(b) = f(bb) + 12 f(Bb), because every homozygote contributes two copies of its allele while every heterozygote contributes one of each. The two allele frequencies must sum to 1 as a sanity check.
- Convert the percentages to fractions. f(BB) = 0.22, f(Bb) = 0.62, f(bb) = 0.16. Sanity check: 0.22 + 0.62 + 0.16 = 1.00.
- Compute p = f(B). p = f(BB) + 12 f(Bb) = 0.22 + 12(0.62). Evaluate 12 × 0.62 = 0.31. So p = 0.22 + 0.31 = 0.53.
- Compute q = f(b). q = f(bb) + 12 f(Bb) = 0.16 + 0.31 = 0.47.
- Sanity check. p + q = 0.53 + 0.47 = 1.00.
- Convert to percentages. p = 53% and q = 47%.
f(B) = p = 0.53 = 53% and f(b) = q = 0.47 = 47%.
Strategic angle. ``Count'' allele copies. Each BB has 2 B's, each Bb has 1 of each, each bb has 2 b's. Divide by 2N (total alleles).
- Imagine a population of 100 individuals. Then 22 BB, 62 Bb, 16 bb. Total alleles = 200.
- Count B alleles. 22 × 2 + 62 × 1 = 44 + 62 = 106. So f(B) = 106/200 = 0.53.
- Count b alleles. 62 × 1 + 16 × 2 = 62 + 32 = 94. So f(b) = 94/200 = 0.47. Check 0.53 + 0.47 = 1.00.
Why this matters. Plugging p and q into Hardy–Weinberg then predicts the genotype frequencies under equilibrium and reveals whether the observed population is in HW equilibrium or evolving.
Allele-counting cross-check. In 100 individuals: 22 BB contribute 44 B, 62 Bb contribute 62 B and 62 b, 16 bb contribute 32 b. Total B = 106, total b = 94 out of 200 alleles, giving p = 0.53 and q = 0.47 –- identical to the algebraic answer, confirming the formula is just bookkeeping.
f(B) = 53%, f(b) = 47%.
Among the five factors that are known to affect Hardy–Weinberg equilibrium, three factors are gene flow, genetic drift and genetic recombination. What are the other two factors?
Concept used. The Hardy–Weinberg equilibrium holds in an idealised population that has no mutation, no natural selection, no gene flow, no genetic drift and random mating (no preferential mating, hence no genetic-recombination bias). Disturb any of these and allele/genotype frequencies shift –- the population evolves. Three of the disturbing forces are listed in the question; the other two are mutation and natural selection.
- List all five forces. 1. Gene flow, 2. Genetic drift, 3. Mutation, 4. Natural selection, 5. Genetic recombination from non-random mating.
- Subtract the three given. Question gives gene flow, genetic drift, genetic recombination. The remaining two are mutation and natural selection.
The remaining two factors are mutation and natural selection.
Quick reading. Five-minus-three = two. Recall the canonical list and subtract.
- Canonical five: gene flow, drift, mutation, selection, recombination.
- Subtract: gene flow, drift, recombination given ⇒ mutation and natural selection remain.
Mnemonic. M S D G R –- Mutation, Selection, Drift, Gene flow, Recombination. The question hides three (G, D, R) and asks for the remaining two (M and S) –- the two most ``Darwinian'' of the five, since mutation supplies the raw material and natural selection imposes the direction.
Mutation and natural selection.
What is founder effect?
Concept used. The founder effect is a special case of genetic drift that occurs when a small number of individuals (``founders'') break away from a large parent population and establish a new colony. By pure chance, the founders carry only a small, possibly non-representative subset of the parent population's alleles. Some alleles that were common in the parent may be absent in the founders; some that were rare may be over-represented. The new population then evolves from this skewed starting point, often becoming markedly different from the parent over a few generations.
- Picture the event. A large parent population on the mainland; a small group (say a dozen people, or a few birds blown off course) reaches a remote island and starts a new population.
- Sampling error. The founders' allele frequencies are a random sample of the parent's; with a small sample, sampling error is large.
- Long-term consequence. The new population may show unusually high frequencies of certain alleles (e.g. rare disease alleles), unusually low diversity, and rapid divergence from the ancestral gene pool.
Founder effect: a form of genetic drift in which a small group of individuals migrates to a new area, carrying a non-representative subset of the parent population's alleles; the new population's gene pool starts markedly different from the ancestral one.
Quick reading. ``Founder'' = the small set of original colonists. ``Effect'' = the resulting skewed gene pool.
- Cause. A few individuals leave the parent population.
- Mechanism. Sampling error in their allele frequencies.
- Outcome. The new colony's gene pool diverges sharply from the parent.
Bottleneck vs. founder. Both are kinds of genetic drift driven by tiny population size. Bottleneck: a once-large population crashes (cheetahs, northern elephant seals), leaving impoverished diversity. Founder: a small subset starts a new population (Amish, Tristan da Cunha). NEET often pairs the two terms and asks which is which.
Genetic-disease lens. Pingelapese in Micronesia show ∼10% incidence of complete colour-blindness (one in ten, vs. one in 30 000 globally) traceable to a single 18th-century cyclone survivor; the Old Order Amish carry abnormally high frequencies of Ellis–van Creveld syndrome, glutaric aciduria and pyruvate kinase deficiency –- each a founder-effect signature still visible today.
A small founding group establishes a new population whose allele frequencies differ from the parent population's, due to sampling error (a sub-type of genetic drift).
Who among the Dryopithecus and Ramapithecus was more man-like?
Concept used. Both Dryopithecus and Ramapithecus are Miocene-age primates (∼15 mya) known from the Siwalik fossil beds. Dryopithecus was more ape-like –- it had long arms, walked in trees and resembled gibbons in proportions. Ramapithecus was more man-like, with smaller canines, a flatter face, jaws that worked more side-to-side (chewing pattern), and probable ground-living habits –- features pointing toward the hominid line.
- Dryopithecus traits. Long forelimbs, tree-dwelling, large canines, V-shaped jaw –- ape-like.
- Ramapithecus traits. Smaller canines, parabolic dental arch (closer to the modern human U-shape), reduced facial prognathism, evidence of ground-living lifestyle –- man-like.
- Compare. Ramapithecus is more hominid in dental and jaw anatomy.
Ramapithecus was the more man-like of the two; it had smaller canines, a more human-like jaw shape and probable ground-dwelling habits, whereas Dryopithecus was more ape-like.
Quick reading. Look at the teeth and jaw. Smaller canines and a flatter, parabolic jaw = more man-like.
- Ramapithecus: small canines, parabolic dentition.
- Dryopithecus: long canines, V-shaped dentition.
Dental signatures in the Siwaliks. Ramapithecus canines were small and the molars heavily worn flat –- the classic plant-grinder pattern that aligns with hominid (human-family) dentition. Dryopithecus retained the long pointed canines and V-shaped (parallel-sided) jaw of generalised apes. Dentition is the single most reliable hominid marker in the Miocene record, and many of these specimens were first described from the Indian Siwalik Hills.
Ramapithecus was the more man-like form.
By what Latin name the first hominid was known?
Concept used. The first known hominid (a primate of the human family Hominidae, exhibiting bipedalism and other human-like traits) was named Homo habilis –- literally ``handy man''. The name refers to its association with the earliest known stone tools (Oldowan tradition) at sites in East Africa. Homo habilis lived approximately 2 mya and had a cranial capacity of ∼650–800 cc, larger than Australopithecus. (Note: some Exemplar reference materials accept the older interpretation pointing to Ramapithecus as the earliest hominid form; on modern phylogenies, however, Homo habilis is the first true member of genus Homo.)
- Define hominid. A member of family Hominidae –- great apes and humans –- traditionally focused on the human lineage.
- First true member of genus Homo. Homo habilis –- the ``handy man'', earliest tool-maker.
- Cite the Latin name. Homo habilis (Leakey, 1964).
The first true hominid of genus Homo is Homo habilis (``handy man'').
Quick reading. ``First hominid'' in the NCERT chronology is the first member of genus Homo, which is Homo habilis.
- Translate the Latin. Homo = human, habilis = handy/skilful.
- Why this matters. Homo habilis is the first member of the human genus to make stone tools.
NCERT wording note. Some Exemplar reference keys accept the older interpretation that Ramapithecus (later reclassified within Sivapithecus) was the first ``hominid'' in a very loose sense. Modern phylogenetics resolves the ambiguity: the first true member of genus Homo is Homo habilis, and that is the safe NEET answer when the question asks for a Latin name.
Homo habilis.
Among Ramapithecus, Australopithecines and Homo habilis –- who probably did not eat meat?
Concept used. Diet of fossil hominids is inferred from dental morphology (size and shape of teeth) and from associated tool/butchering evidence. Ramapithecus (∼15 mya) had relatively small canines, broad flat molars and powerful chewing musculature –- the classic signature of a herbivore (probably a seed and fruit eater). Australopithecines and Homo habilis, on the other hand, show evidence (smaller jaws, scavenger bite marks on bones at their sites, stone tools used to cut flesh) of meat consumption alongside plant food.
- Ramapithecus. Heavy flat molars, small canines, no stone tools known –- diet was almost entirely plant-based. Did not eat meat.
- Australopithecines. Stone tools at Australopithecus sites and butchered bones suggest some meat eating.
- Homo habilis. Oldowan stone tools and clear butchering marks on antelope bones indicate active meat eating (probably scavenged at first).
Ramapithecus probably did not eat meat; its dentition is that of a strict herbivore (seed/fruit eater).
Quick reading. Look at the teeth. Flat molars, no tools ⇒ herbivore. Ramapithecus fits.
- Ramapithecus. Flat molars, no tools ⇒ herbivore.
- Australopithecines and Homo habilis. Tools + butchered bones ⇒ some meat.
Tool-use evidence. Oldowan stone flakes at Olduvai Gorge (Tanzania) date to 2.5 Ma and are found alongside antelope long-bones with cut-mark patterns produced by stone edges, not by carnivore teeth –- the smoking-gun evidence that Homo habilis butchered carcasses. No tools and no butchered bones accompany Ramapithecus sites, reinforcing the herbivore inference.
Brain plus tools. The earliest Oldowan tools (2.5–1.7 Ma) needed cognitive planning, motor coordination and the social transmission of know-how –- traits absent in Ramapithecus. Hominid carnivory and tool-use co-evolved, each enabling the brain expansion that defines the genus Homo.
Ramapithecus (herbivorous).
Short Answer Type Questions
Louis Pasteur's experiments, if you recall, proved that life can arise from only pre-existing life. Can we correct this as life evolves from pre-existent life or otherwise we will never answer the question as to how the first forms of life arose? Comment.
Concept used. Pasteur's swan-necked-flask experiments (1861) disproved spontaneous generation in present-day Earth conditions –- meat broth exposed to airborne dust grew microbes, but broth in a curved-neck flask that trapped dust stayed sterile. The conclusion was: under modern conditions, life arises only from pre-existing life (biogenesis). But this leaves unsolved how the first life ever arose. The accepted scientific correction is the Oparin–Haldane chemical-evolution hypothesis: the very first living forms arose by chemical evolution from non-living organic molecules under the very different reducing-atmosphere conditions of the primitive Earth.
- What Pasteur actually proved. Under present atmospheric and biotic conditions, life arises only from existing life. Pasteur's flasks were stocked with rich nutrient broth, exposed to ordinary modern air; nothing grew unless airborne microbes reached them.
- The gap Pasteur's result leaves. His finding cannot tell us what happened 3.8–4 bya on a sterile Earth with no living microbes and a very different reducing atmosphere. Restricting biology to ``life from life'' would push the origin of the first life out of the realm of science.
- The correction. Rephrase Pasteur as: contemporary life evolves from pre-existing life. The very first life arose from non-living organic matter through chemical evolution –- the Oparin–Haldane / Miller–Urey scenario. This restores a scientific path for the origin question.
- Conclusion. Yes, the statement should be corrected to ``life evolves from pre-existing life''; otherwise the origin of the first life becomes scientifically un-askable.
Pasteur's law applies to existing life under modern conditions. To explain the very first life, we accept that life evolves from pre-existent life now, but originally arose from non-living organic molecules under the primitive Earth's reducing atmosphere (Oparin–Haldane chemical evolution).
Strategic angle. Separate the question into two time-frames: today and the very first day.
- Today. Pasteur is right –- biogenesis: life from life.
- The first day (3.8 bya). Biogenesis cannot start the chain; chemical evolution must.
- Bridge. The corrected slogan ``life evolves from pre-existent life'' allows continuity once life exists, while Oparin–Haldane explains how it got started.
Why this matters. Without this two-step view, biology faces an infinite-regress (where did the first life's parent come from?) –- acknowledging chemical evolution closes the regress.
Time-scale frame. On modern Earth, biotic competition is so intense that any prebiotic ``soup'' would be eaten by existing microbes before it could organise into a new life –- Pasteur is right here, now. On the pre-biotic Earth there were no microbes to eat anything, so chemistry had a free hand for ∼500 million years before the Last Universal Common Ancestor emerged ∼3.8 bya. Two regimes, two correct answers –- the apparent contradiction dissolves with time-scale awareness.
Yes, correct it to ``life evolves from pre-existent life'', and add chemical evolution to account for the very first life.
The scientists believe that evolution is gradual. But extinction, part of evolutionary story, are 'sudden' and 'abrupt' and also group-specific. Comment whether a natural disaster can be the cause for extinction of species.
Concept used. Evolution is typically gradual, proceeding by small heritable changes accumulating over many generations. Mass extinction, however, is the relatively sudden loss of a large fraction of species across a short geological interval, often triggered by a single global event. Five major mass extinctions are recognised, the most famous being the end-Cretaceous (K-Pg) event 65 mya in which the non-avian dinosaurs (and many marine groups) disappeared –- attributed to a massive asteroid impact in Yucat'an, possibly compounded by Deccan volcanism. A natural disaster of sufficient scale (asteroid impact, super-volcano, sudden glaciation, large sea-level change) can absolutely cause group-specific extinction by destroying a habitat type faster than its specialists can adapt or migrate.
- Mechanism of disaster-driven extinction. A natural disaster (asteroid, volcanism, sudden climate shift, large fire) abruptly alters environmental parameters (temperature, light, available habitat). Species lacking the genetic variation to tolerate the new conditions die out.
- Why it appears sudden in the fossil record. The change occurs over thousands of years (geologically fast), so in sedimentary layers it looks like an abrupt cliff.
- Why it appears group-specific. Different species have different ecological tolerances. A cold spell wipes out tropical specialists more than temperate ones; an asteroid winter wipes out large diurnal animals (dinosaurs) more than small burrowers (mammals).
- Example. K-Pg boundary 65 mya: the Chicxulub impact ejected sulphate aerosols, blocked sunlight for years, collapsed photosynthesis. Large herbivores starved, then their predators. Small endotherms with diverse diets survived.
Yes, a major natural disaster (asteroid, super-volcanism, abrupt climate change) can cause sudden, group-specific extinction by altering the environment faster than affected species can adapt –- the K-Pg event is the textbook example.
Strategic angle. Reconcile two facts: evolution is slow, extinction can be fast. A natural disaster supplies the speed gap.
- Slow background. Normal evolution and ``background'' extinction.
- Disaster spike. A single event rewrites environmental rules globally in years to centuries.
- Selective loss. Groups whose tolerance breaks first disappear; the spike looks group-specific.
Why this matters. Today's biodiversity crisis (the proposed ``sixth extinction'') is essentially a slow-motion human-caused natural disaster –- same logic, different agent.
Six extinctions in the rock. End-Ordovician (∼444 Ma, glaciation), late Devonian (∼375 Ma, anoxia), end-Permian (252 Ma, ``Great Dying'', super-volcanism, ∼96% marine species lost), end-Triassic (201 Ma, volcanic CO2), end-Cretaceous (65 Ma, Chicxulub asteroid plus Deccan Traps) –- and the ongoing Anthropocene loss, driven by the same logical agent: a fast environmental shift outrunning species' adaptive capacity.
Modern parallels. The current Anthropocene-extinction wave is running at ∼100–1000 times background extinction rate, driven by habitat destruction, climate change and invasive species –- a slow-motion natural disaster of human origin. Mammals, amphibians and corals are losing species fastest, exactly mirroring the group-specificity of K-Pg in a different lineage profile. The lesson is the same: when environmental change outruns adaptive response, biodiversity collapses non-randomly.
Yes; natural disasters can cause sudden, selective extinctions by exceeding species' adaptive limits.
Why is nascent oxygen supposed to be toxic to aerobic life forms?
Concept used. Nascent oxygen refers to atomic or highly reactive forms of oxygen –- single O atoms, superoxide radical O2^.-, hydroxyl radical OH^., hydrogen peroxide H2O2 –- collectively called reactive oxygen species (ROS). Unlike molecular O2, these species have unpaired electrons or weak O-O bonds and are extremely reactive: they oxidise lipids (membrane damage), proteins (loss of enzyme function), and DNA (strand breaks, base modifications). Aerobic organisms therefore evolve enzymes (superoxide dismutase, catalase, glutathione peroxidase) to neutralise ROS; without them, even essential aerobes are poisoned by their own oxygen metabolism.
- Why ROS are reactive. They carry unpaired electrons (O2^.-, OH^.) or unstable O-O bonds (H2O2). To reach a stable closed-shell configuration, they snatch electrons from organic molecules.
- What they damage. Membrane lipids (peroxidation chain reactions), enzymes (oxidising cysteine and methionine), DNA (8-oxoguanine, strand breaks, base loss leading to mutation).
- Why aerobic cells survive at all. They possess defensive enzymes –- catalase converts H2O2 to H2O and O2; superoxide dismutase converts O2^.- to H2O2; glutathione peroxidase reduces peroxides at the expense of GSH.
- Without these enzymes. The cell would be killed by its own oxidative metabolism. Nascent oxygen is therefore intrinsically toxic; aerobes thrive only because they detoxify it continuously.
Nascent oxygen (atomic O, O2^.-, OH^., H2O2) is chemically very reactive: it oxidises lipids, proteins and DNA, killing unprotected cells. Aerobes survive by deploying detoxifying enzymes (catalase, SOD, glutathione peroxidase).
Strategic angle. Atomic oxygen is electronically unstable –- it wants to grab an electron from something. ``Something'' is the cell's biomolecules.
- Free radicals. O^., O2^.-, OH^. have unpaired electrons.
- Reaction with cell. They attack lipids (membrane), proteins (enzymes) and nucleic acids (DNA).
- Result. Cell death unless ROS scavenger enzymes intervene.
Why this matters. The oxidative-stress theory of ageing pins gradual cellular decline on cumulative ROS damage –- the same nascent oxygen the question asks about.
Cellular cost of aerobic life. Mitochondrial electron transport leaks ∼1–2% of consumed O2 as superoxide; a human burns ∼400 L O2/day, so ∼5 L is shunted through ROS pathways. Without catalase, SOD and GSH peroxidase, this would shred every protein and lipid in the body within hours. The same ROS pressure is thought to have delayed the appearance of multicellular aerobes by ∼1 Gyr after the Great Oxygenation Event.
ROS as evolutionary throttle. The first cyanobacteria appeared ∼2.7 bya; they took ∼200 myr to oxygenate the oceans and another ∼1 Gyr before complex multicellular life appeared. The lag reflects how long it took organisms to evolve the SOD/catalase/peroxidase tool-kit that lets cells survive the same oxygen they now need. Nascent oxygen was therefore the chemical that both enabled (via efficient ATP yield) and gated (via ROS toxicity) the rise of complex life.
Atomic/radical oxygen is highly reactive and damages lipids, proteins and DNA; aerobic cells need detoxifying enzymes to survive their own oxygen.
While creation and presence of variation is directionless, natural selection is directional as it is in the context of adaptation. Comment.
Concept used. The two steps of Darwinian evolution have opposite character. Step 1: generation of variation (mutation, recombination) is random and undirected –- mutations occur wherever DNA chemistry slips, regardless of usefulness. Step 2: natural selection is directional –- the environment preferentially preserves variants that improve survival and reproduction in that environment, so trait distributions shift toward better-adapted forms over generations. This combination of random variation with directional selection is the core of neo-Darwinism.
- Random variation. Mutations arise as chemical accidents. They have no foresight; some will be neutral, some harmful, a few beneficial. The variation pool is therefore omnidirectional.
- Directional selection. Once variation exists, the environment is a strict filter. In hot dry climates, water-saving variants leave more offspring than water-wasting variants. The selection vector points consistently toward better-adapted forms, which is why we say it is directional.
- Together. Random variation supplies the raw material; directional selection imposes the direction. Adaptation emerges as a statistical outcome of these two steps over many generations.
Mutation and recombination supply variation randomly (no direction); natural selection then propagates variants that fit the environment, giving evolution a clear adaptive direction.
Strategic angle. Two-step engine: blind shuffle, then directional filter.
- Step 1. Variation is generated by random mutation and recombination.
- Step 2. Selection filters variants by their fitness in a specific environment.
- Net result. Populations adapt –- not because variation ``tries'' to adapt, but because the environment keeps reposting survivors.
Why this matters. Confusing the two steps is the Lamarckian error: believing variation itself is goal-directed. NEET often tests this exact distinction.
Cement of neo-Darwinism. The two-step picture –- random variation, directional selection –- is what separates evolutionary biology from teleology. Lamarck assumed variation itself ``aimed'' at adaptation (giraffe necks stretched on purpose); Darwin separated the two steps; the Modern Synthesis (1930s–40s) supplied the molecular basis (mutation + meiotic recombination as the random-variation engine). Mixing the steps is the most common Lamarckian relapse on NEET.
Variation is random (undirected); selection is directional. The combination produces adaptive evolution.
The evolutionary story of moths in England during industrialisation reveals, that 'evolution is apparently reversible'. Clarify this statement.
Concept used. The Biston betularia peppered-moth story is the classic short-time-scale example of directional natural selection. Pre-Industrial Revolution: tree bark in England was pale because lichens covered it; pale moths were camouflaged and dark variants were eaten by birds, so pale moths dominated (>99% of the population). During the Industrial Revolution: soot blackened bark and killed lichens; dark moths were now camouflaged, pale moths were eaten, and dark moths rose to >90%. After Clean Air Act (1956): pollution fell, lichens recovered, bark became pale again, and pale moths rose once more. The population's mean colour shifted dark, then back to pale –- evolution apparently reversed direction.
- Pre-industrial state. Pale bark, pale lichens. Pale moths camouflaged; dark moths eaten. Frequency of dark moths ∼1%.
- Industrial state. Soot blackens bark, kills lichens. Now dark moths camouflaged; pale moths conspicuous and eaten. Frequency of dark moths rises to ∼95% in heavily polluted cities (Manchester) within ∼50 years.
- Post-clean-air state. Pollution falls, lichens recover, bark returns to pale. Selection reverses: dark moths eaten, pale ones survive. Pale moths rise again.
- Interpretation of ``reversible''. Both alleles (carbonaria dark, typica pale) remained in the gene pool throughout. Selection simply reversed direction, swinging the frequencies back and forth. The genes were never lost –- only the frequencies oscillated. Evolution looks reversible only in the sense that the same trait can be re-favoured if the environment swings back.
In Biston betularia, the dark/pale frequency rose under industrial pollution and fell back after clean air –- because both alleles remained in the gene pool, selection simply swung in opposite directions, making evolution apparently reversible.
Strategic angle. Track the dominant phenotype before, during and after pollution.
- Before: pale moths dominate. During: dark moths dominate. After clean air: pale moths rise again.
- Why it reversed. The environment reversed; selection reversed; the pre-existing alleles re-emerged.
- What ``apparently'' means. The alleles never died out, so the frequency could swing back. True reversal of lost evolution (Dollo's law) is not possible.
Why this matters. The moth case is the most-quoted real-time proof of natural selection –- and a reminder that selection has no memory.
The numbers. Pre-1850 dark-morph frequency in rural Dorset was below 2%; by 1900 in industrial Manchester it was above 95%; after the 1956 Clean Air Act dark frequency in Manchester fell back to ∼10% by 2000. Each transition tracks SO2 and soot deposition almost in step –- making Biston betularia the best-documented case of natural selection in any wild animal.
Why ``apparently'' matters. If the dark allele had been lost outright during pre-industrial times (frequency = 0, not ∼1%), the population could not have responded to industrial soot at all, because mutation rates (∼ 10-6 per generation) are too slow to regenerate the allele in ∼50 years. The episode therefore also illustrates that standing genetic variation is what makes rapid evolution possible –- a key Hardy–Weinberg implication. Dollo's law (true loss is irreversible) remains intact; what reversed here were frequencies, not the alleles themselves.
Pollution rise → dark moths rise. Clean air → pale moths return. Both alleles persist, so the frequencies swing back; evolution looks reversible but the genes were never lost.
Comment on the statement that ``evolution and natural selection are end result or consequence of some other processes but themselves are not processes''.
Concept used. Evolution (change in allele/genotype frequencies across generations) and natural selection (differential reproductive success of variants) are not stand-alone processes that ``do'' something on their own. They are the outcomes of more fundamental biological processes: mutation and recombination generate heritable variation; reproduction hands variants to the next generation; the environment filters which variants leave more offspring. Without these underlying processes, neither selection nor evolution would occur.
- Identify the underlying processes. Mutation (DNA chemistry), meiotic recombination (chromosomal shuffle), reproduction with heritability, and environmental interaction (predation, climate, food).
- See selection as a consequence. ``Differential reproductive success'' is just a tally –- it emerges as the consequence of variants meeting the environment. The environment is not selecting consciously; it just imposes survival challenges.
- See evolution as a consequence. ``Change in allele frequencies'' is the statistical result of selection, mutation, drift, gene flow operating across generations.
- Why the statement is correct. Evolution and selection have no machinery of their own; they are emergent statistical descriptions of what variation and the environment together produce.
Yes –- evolution and natural selection are not active processes but emergent statistical outcomes of mutation, recombination, reproduction and environmental interaction. They describe the result, not the mechanism.
Strategic angle. ``Process'' implies active machinery. Selection and evolution have no machinery; they are descriptions of outcomes.
- Active processes. Mutation chemistry, recombination biology, reproduction physiology, environment dynamics.
- Passive descriptions. ``Natural selection'' = the tally of who reproduces. ``Evolution'' = the running record of allele frequencies.
- Therefore. The statement is correct: selection and evolution are consequences, not processes.
Why this matters. This view dissolves popular ``selection is a designer'' misconceptions –- selection has no agency.
Avoiding the agency trap. ``Selection chose'' is shorthand. Strictly, there is no chooser; there are only individuals reproducing at different rates because their heritable traits differ in fit to the environment. Allele frequencies then shift as a statistical outcome. Removing the agency language sharpens every NEET answer that touches on natural selection.
Correct: evolution and natural selection are outcomes of mutation, recombination, reproduction and environmental interaction; they have no independent machinery.
State and explain any three factors affecting allele frequency in populations.
Concept used. The Hardy–Weinberg principle states that allele frequencies stay constant generation to generation if a population has no mutation, no selection, no migration, no drift and random mating. Any of the five evolutionary forces disturbs this equilibrium and changes allele frequency. Three of the most important are mutation, natural selection and genetic drift.
- Mutation. Random heritable changes in DNA convert one allele to another (A → a or vice versa). Even at low per- generation rates (10-9 to 10-5 per site per generation), mutation is the ultimate source of new alleles and gradually shifts allele frequencies. It is also the only force that introduces new genetic material; the others merely rearrange existing alleles.
- Natural selection. If allele A confers higher survival or reproductive success than allele a, A-bearing individuals produce more offspring. The frequency of A rises generation by generation. Selection is the directional force –- it pushes frequencies toward the locally fittest variant.
- Genetic drift. In a finite population, the alleles passed to the next generation are a small random sample of the parent gene pool. Sampling error causes allele frequencies to drift up or down by chance, independent of fitness. The smaller the population, the stronger drift becomes. In tiny populations, rare alleles can be lost entirely, and common ones can become fixed –- with no help from selection.
- Two additional forces (not asked but worth mentioning). Gene flow (migration of individuals between populations carries alleles in or out) and non-random mating (e.g. inbreeding raises homozygote frequency without changing allele frequency immediately, but biases mating choices over time affect allele frequencies via selection differentials).
Three factors that change allele frequency: mutation (new alleles), natural selection (differential reproductive success), genetic drift (random sampling in finite populations).
Strategic angle. Each force shifts allele frequency by a characteristic mechanism: create, filter, or shuffle by chance.
- Create: mutation. New alleles appear.
- Filter: natural selection. Beneficial alleles spread.
- Shuffle by chance: genetic drift. Random sampling moves frequencies up or down.
Why this matters. Each force has a distinct signature in real populations. Mutation gives steady low-rate change; selection gives directional sweeps; drift gives fluctuations whose size scales as 1/√N.
Quick scaling guides. Drift effect ∝ 1/√N –- weak in millions, devastating in dozens. Mutation rate ≈ 10-8 per nucleotide per generation in humans –- slow but steady. Selection coefficient s as small as 10-3 can fix an allele in ∼10 000 generations –- powerful given enough time. Each force has a characteristic strength that NEET MCQs test with magnitude comparisons.
Worked illustration. Take a small island population of 50 lizards where allele A starts at 0.5 and confers a 5% survival advantage. Selection alone raises f(A) by ∼0.013 per generation; drift fluctuates it by ∼ 1/√100 = 0.1 per generation; if 5% migrate per generation from a mainland fixed for a, gene flow drags f(A) down by 0.025 per generation. The three forces compete; whichever has the largest signed effect wins. This single sketch summarises every NEET-level question on allele-frequency change.
Mutation, natural selection and genetic drift are three independent forces that shift allele frequencies.
Gene flow occurs through generations. Gene flow can occur across language barriers in humans. If we have a technique of measuring specific allele frequencies in different population of the world, can we not predict human migratory patterns in pre-history and history? Do you agree or disagree? Provide explanation to your answer.
Concept used. Gene flow is the movement of alleles between populations through migration and interbreeding. When a migrating group settles in a new area and intermarries with the resident population, it imports its own alleles and exports residents' alleles. Allele frequencies, especially of distinctive variants (Y-chromosome haplogroups, mitochondrial DNA lineages, specific SNPs), thus leave a chemical record of ancient migration. Modern population genetics reads this record to reconstruct human migratory history –- ``genetic archaeology''.
- Agree with the statement. Yes, allele frequencies in modern populations are a fossil of ancient migration.
- Reason 1: distinctive markers. Some alleles arose by single mutations in one ancestral region (e.g. certain Y-chromosome haplogroups arose in Africa or Central Asia). Their spatial distribution today maps the spread of their carriers.
- Reason 2: gradient analysis. A gradient of allele frequency from a putative origin to a frontier indicates a wave-of-advance migration. The lactase-persistence allele (LCT/MCM6) is a textbook case: frequency >70% in northern Europe, <10% in East Asia, mapping the spread of cattle-herding populations.
- Reason 3: mitochondrial DNA and Y-chromosome trees. Maternally inherited mtDNA and paternally inherited Y-DNA do not recombine, so their lineages can be traced like family trees across populations –- yielding the ``Out of Africa'' model and the populating of Eurasia, Americas and Australia.
- Concrete result. Genetic studies confirm independent migrations across the Bering land-bridge (peopling of the Americas, ∼15 kya), the Bantu expansion across sub-Saharan Africa, the Austronesian expansion across the Pacific, the Indo-European spread across Eurasia. All were inferred from allele-frequency distributions.
Agree. Allele frequencies (especially Y-chromosome haplogroups, mtDNA lineages and distinctive SNPs) record ancient migrations. Population-genetics analyses of these data have reconstructed Out-of-Africa dispersal, peopling of the Americas, Bantu, Austronesian and Indo-European migrations.
Strategic angle. Treat each distinctive allele as a postal mark on a parcel: it was made in one place at one time, and it travelled wherever its carriers travelled.
- Origin and spread. Mutation arises in one population at time T; carriers later migrate, depositing the allele in new regions.
- Frequency map. Plot the allele's frequency on a world map. A gradient pointing away from the origin reveals the migration direction.
- Combine many alleles. With dozens of independent markers, the routes triangulate sharply.
Why this matters. The 1000 Genomes Project and the Genographic Project have explicitly used allele-frequency data to reconstruct human prehistory.
Routes already mapped. The peopling of the Americas via the Bering land bridge ∼15–25 ka, the Bantu agricultural expansion across sub-Saharan Africa from ∼3 ka, the Austronesian seafarer dispersal from Taiwan across the Pacific from ∼4 ka, and the Indo-European Steppe-pastoralist expansion across Eurasia ∼5 ka –- every one was reconstructed first from allele-frequency gradients, then confirmed by archaeology and linguistics.
Yes, allele frequencies do encode ancient human migrations and allow us to reconstruct prehistoric movements.
How do you express the meaning of words like race, breed, cultivars or variety?
Concept used. Race, breed, cultivar and variety are all infraspecific (below species) labels for groups of organisms within a single species that share heritable distinctive traits. The four terms are used in slightly different contexts but share a common biological meaning: a subset of a species with a recognisable genetic and phenotypic identity, maintained either by geographical isolation, by reproductive isolation, or by deliberate human breeding.
- Race. A genetically distinct subset of a wild species adapted to a particular geographical area; closer to ``ecotype'' or ``subspecies'' in modern usage. Used historically in humans, though modern genetics finds human ``races'' biologically poorly defined.
- Breed. A genetically distinct subset of a domesticated animal species, produced by selective breeding for traits such as milk yield, wool quality or temperament. Examples: Sahiwal cattle, Murrah buffalo, Labrador dog.
- Cultivar. A ``cultivated variety'' –- a genetically distinct subset of a plant species developed and maintained by deliberate breeding for desirable agronomic traits (high yield, disease resistance). Examples: Sonalika wheat, Pusa Basmati rice.
- Variety. The general botanical term for any infraspecific plant taxon; in cultivated species ``variety'' overlaps with cultivar. In wild species, ``variety'' is a named natural subgroup below subspecies.
All four terms label genetically distinct subsets within a single species. Race = geographical/ecological subset (wild). Breed = human-bred subset of domesticated animals. Cultivar = human-bred subset of cultivated plants. Variety = general infraspecific botanical group.
Strategic angle. Two axes distinguish the four terms: domain (animal vs. plant) and origin (wild vs. human-bred).
- Race: wild, often human ancestry usage.
- Breed: domesticated animals, human-bred.
- Cultivar: cultivated plants, human-bred.
- Variety: general botanical infraspecific category, wild or cultivated.
Why this matters. Indian breeding programmes use exactly these labels: ``Sahiwal breed'' (cattle), ``Sonalika cultivar'' (wheat), ``Indica variety'' (rice), all within a single species each.
Indian taxonomy examples. Bos indicus race ``Sahiwal'' (cattle breed, milk-yield trait fixed); Triticum aestivum cultivar ``Sonalika'' (wheat, semi-dwarf high-yield); Oryza sativa variety ``IR8'' (rice, Green-Revolution short stature); Cocos nucifera variety ``Chowghat Orange Dwarf'' (coconut, oil yield) –- each is an infraspecific group with a name that signals its origin.
Why naming matters. The species Bos indicus contains ∼30 named cattle breeds in India alone; the species Triticum aestivum contains hundreds of named cultivars worldwide. Without infraspecific labels, breeders could not track which heritable subgroup carries which trait. The four terms therefore have legal weight under the Plant Variety Protection Act (India, 2001) and the International Convention for the Protection of New Varieties of Plants.
Race, breed, cultivar and variety are all infraspecific labels for genetically distinct subgroups within a species, differing by domain (animal/plant) and origin (wild/human-bred).
When we say ``survival of the fittest'', does it mean that
(a) those which are fit only survive, or
(b) those that survive are called fit?
Comment.
Concept used. ``Survival of the fittest'' (Herbert Spencer's phrase, later adopted by Darwin) sounds tautological if read as ``the survivors are by definition fit''. The precise neo-Darwinian meaning is that organisms with heritable traits that better suit the prevailing environment leave more offspring than less-suited organisms; fitness is measured by relative reproductive success, not by simply surviving the moment. Reading (a) is correct only with the caveat that ``fit'' means adapted in advance (carrying useful heritable variation) –- not ``fortunately alive''. Reading (b) is a tautology and is therefore not the scientifically meaningful interpretation.
- Interpretation (a). Fitness is a heritable property existing before the environmental test. The fit individuals carry alleles that confer better survival or reproduction in that environment. They then survive and reproduce because they were fit. This is the Darwinian reading.
- Interpretation (b). Survivors are post-hoc labelled ``fit''. This is circular: fitness = survival, survival explains fitness, fitness explains survival –- a closed loop with no explanatory power. This reading is rejected.
- Resolve the apparent tautology. Fitness is best defined operationally as relative reproductive success in a given environment. The trait responsible for high fitness can be independently identified (camouflage, drought tolerance, etc.). With this operational definition, ``the fittest survive'' is a substantive empirical claim, not a tautology.
Reading (a) is correct when ``fit'' means ``carrying heritable traits suited to the environment''; those individuals leave more offspring. Reading (b) is a circular tautology and is rejected.
Strategic angle. Decide whether ``fitness'' is a cause or a description, then pick (a) or (b).
- If fitness is causal (heritable trait → better reproduction), then ``the fit survive'' is meaningful science. Pick (a).
- If fitness is descriptive (survivors = fit by definition), the statement is tautology. Reject (b).
Why this matters. Modern fitness measurements (lifetime reproductive output, finch-beak data, drug-resistant bacterial growth curves) all use the causal definition –- exactly Darwin's intent.
Operational rescue. Modern biology rescues the slogan by measuring fitness as ``lifetime number of offspring that themselves reproduce'' –- a quantity independent of any prior survival claim. Once fitness has this independent yardstick (Daphne Major finch beak depth predicting drought-year survival is the canonical example), the statement ``fittest survive'' becomes a testable empirical claim rather than a circular definition.
Why the resolution matters. Karl Popper once dismissed Darwinism as untestable for exactly this circularity reason. Modern evolutionary biology answers by tying fitness to specific, measurable traits before testing whether high-fitness individuals indeed leave more offspring. Once fitness is operationalised, the tautology dissolves and the prediction becomes falsifiable.
Reading (a) is correct: ``fit'' = pre-existing adaptation; survival is the consequence, not the definition.
Enumerate three most characteristic criteria for designating a Mendelian population.
Concept used. A Mendelian population is the unit of microevolutionary analysis –- a group of sexually reproducing, interbreeding individuals of a single species that share a common gene pool. It is the population in which Mendel's laws of segregation and independent assortment, and the Hardy–Weinberg principle, hold. To qualify as a Mendelian population, a group must satisfy three characteristic criteria.
- Criterion 1: members of a single species. All individuals must belong to the same species so that they are reproductively compatible and can exchange genes through mating.
- Criterion 2: sexual reproduction with interbreeding. The members must reproduce sexually and actually interbreed (or be able to interbreed). Interbreeding is what creates and maintains a shared gene pool.
- Criterion 3: shared gene pool with continuous gene flow among members. Alleles must circulate freely within the group. This is what makes Hardy–Weinberg analysis applicable and what distinguishes a Mendelian population from a collection of unrelated individuals.
- (Implicit fourth criterion.) Geographical contiguity often follows from criterion 3 –- individuals close enough in space to mate frequently –- but it is not always listed separately.
Three characteristic criteria: (i) all members belong to a single species; (ii) members reproduce sexually and interbreed; (iii) they share a common gene pool maintained by continuous gene flow within the group.
Strategic angle. The unit is defined by who can pass alleles to whom. That fixes the criteria.
- One species. Otherwise alleles cannot flow.
- Sexual interbreeding. Otherwise alleles cannot mix.
- Shared gene pool. The cumulative consequence of the first two.
Why this matters. Population geneticists define populations operationally using exactly these three criteria when sampling for allele-frequency studies.
Boundary cases. Allopatric populations of the same species are not a single Mendelian population because gene flow is interrupted; hybrid swarms across a species boundary may not be a Mendelian population because reproductive compatibility breaks down; an asexual clone is not a Mendelian population because Mendel's laws of segregation and independent assortment do not apply. The three criteria define the unit cleanly.
Same species, sexual interbreeding, shared gene pool with free gene flow.
``Migration may enhance or blurr the effects of selection''. Comment.
Concept used. Migration (or gene flow) is the movement of alleles between populations carried by migrating individuals who breed in the new population. Its effect on natural selection depends on which alleles arrive. If incoming alleles match the locally favoured ones, migration enhances selection by adding more fit alleles than the local source rate. If incoming alleles are the locally disfavoured ones, migration counteracts selection by constantly importing maladaptive alleles –- the local population cannot quickly diverge from the parent because gene flow keeps re-mixing its gene pool. Migration is therefore a homogenising force, but it can either accelerate or blunt selection depending on direction.
- Case 1: migration enhances selection. Suppose local environment favours allele A and a few migrants arrive carrying more A than local frequency. Local f(A) rises faster than by selection alone. Selection's effect is amplified.
- Case 2: migration blurs selection. Suppose local environment favours A but migrants keep arriving from a parent population that has high f(a). Each generation, selection raises f(A) while immigration imports a. The two forces cancel, allele frequencies plateau below what pure selection would produce. The local population cannot diverge.
- Empirical consequence. Highly migratory species (oceanic fish, migratory birds) show less local adaptation than sedentary species –- because gene flow keeps blurring local selection. Island populations of the same species often show dramatic local adaptation because gene flow from the mainland is weak.
Migration carries alleles between populations: if it imports alleles favoured locally, it enhances selection; if it imports alleles disfavoured locally, it blurs selection by constantly re-mixing the gene pool. Net effect depends on direction and rate.
Strategic angle. Sign of gene flow vs. sign of selection. Same sign = enhance; opposite sign = blur.
- Same direction. Gene flow imports locally favoured alleles. Selection plus gene flow accelerate divergence in the favoured direction.
- Opposite direction. Gene flow imports locally disfavoured alleles. Selection has to spend effort just countering immigration; local adaptation is held back.
Why this matters. Modern conservation genetics uses this insight: restoring corridors between fragmented populations boosts gene flow –- good for genetic diversity, but potentially blurring local adaptation.
Gene-flow magnitude. As little as one migrant per generation (Nm = 1) is enough to keep two populations from diverging genetically –- Sewall Wright's classical result. That is why island populations a few hundred kilometres offshore can speciate (low m), while continental populations rarely do (high m). The same logic informs corridor design in Indian tiger-reserve conservation.
Conservation flip side. Re-introducing animals from one isolated population into another (genetic rescue) deliberately uses migration to enhance selection for heterozygosity in inbred populations –- as done with Florida panthers (Texas cougar introduction, 1995) and with Indian Asiatic lions in Gir. The same biological lever has very different conservation purposes depending on how it is applied.
Migration enhances selection when it imports locally favoured alleles, and blurs selection when it imports locally disfavoured ones.
Long Answer Type Questions
Name the law that states that the sum of allelic frequencies in a population remains constant. What are the five factors that influence these values?
Concept used. The law that states that allele and genotype frequencies in a large, randomly mating, isolated population remain constant generation after generation is the Hardy–Weinberg principle (also called the Hardy–Weinberg law of genetic equilibrium), formulated independently by G. H. Hardy (a British mathematician) and Wilhelm Weinberg (a German physician) in 1908. For a gene with two alleles A (frequency p) and a (frequency q), p + q = 1, p2 + 2pq + q2 = 1, where p2, 2pq and q2 are the expected frequencies of genotypes AA, Aa and aa respectively. The principle holds only under five idealised assumptions; violation of any one of them causes the allele frequencies to change –- i.e. the population evolves.
- State the principle precisely. In an idealised population, p and q remain constant through generations, so the population is in genetic equilibrium. Deviations between observed and Hardy–Weinberg-predicted genotype frequencies are evidence of evolution at the locus.
- Factor 1 –- Gene flow (migration). Movement of individuals between populations exports and imports alleles. A few migrants per generation can permanently shift allele frequencies; large gene flow homogenises populations.
- Factor 2 –- Genetic drift. In a finite population, the alleles passed to the next generation are a random sample of the parent gene pool. Sampling error causes random fluctuations in p and q; the smaller the population, the stronger the drift. Special cases include the bottleneck effect and the founder effect.
- Factor 3 –- Mutation. Random heritable DNA changes convert one allele to another (e.g. A → a). Mutation rates are low (∼ 10-9 to 10-5 per site per generation) but steady, and mutation is the ultimate source of new alleles.
- Factor 4 –- Genetic recombination. Crossing-over and independent assortment in meiosis reshuffle alleles between chromosomes, creating new gametic combinations. This generates new multilocus genotype frequencies even when single-locus allele frequencies are unchanged.
- Factor 5 –- Natural selection. Differential survival and reproduction of variants. If allele A raises fitness, f(A) rises; if A lowers fitness, f(A) falls. Selection is the directional force.
- Summary table.
tabularlp0.55
Force & Type of change in allele frequency
Gene flow & Homogenising between populations
Genetic drift & Random fluctuation (stronger in small populations)
Mutation & Slow but creates new alleles
Recombination & Reshuffles multilocus combinations
Natural selection & Directional, environment-driven
tabular
The Hardy–Weinberg law (p + q = 1, p2 + 2pq + q2 = 1). Five factors that influence allele frequencies: mutation, natural selection, genetic drift, gene flow (migration) and genetic recombination.
Strategic angle. One law, one equation, five forces.
- Name and equation. Hardy–Weinberg principle: p2 + 2pq + q2 = 1 with p + q = 1.
- Five forces (M S D G R). Mutation, selection, drift, gene flow, recombination.
- Interpretation. Any deviation between observed genotype frequencies and HW prediction is a footprint of one of these five.
Why this matters. Real-population studies of sickle-cell, CCR5, lactase-persistence, MHC alleles all measure HW deviation to detect selection in action.
Equilibrium as null model. Hardy–Weinberg is biology's null hypothesis: under the five idealised conditions, genotype frequencies are predictable from allele frequencies alone. Whenever observed counts differ from p2 : 2pq : q2 in a real population, at least one assumption is violated –- and population geneticists can usually identify which (selection by fitness-trait correlation, drift by population-size estimate, gene flow by migration data). The principle is also the launching pad for modern GWAS analyses, where deviation from HW is the very first quality-control filter on every genotyped SNP.
Calibrating the principle to data. A worked NEET example: in a sample of 1000 humans, if 360 show the MN heterozygote, the calculated 2pq = 0.36, so pq = 0.18, giving p, q ≈ 0.55, 0.45 (or vice-versa). Hardy–Weinberg then predicts MM ≈ 30.3% and NN ≈ 20.3%. An observed deviation of ∼5 percentage points in either homozygote class is enough to suggest selection or non-random mating at the locus. This forensic use of the equation underpins almost every population-genetics paper since the 1960s.
Why all five forces matter together. Each force has a distinct signature: mutation creates new alleles at rate μ ∼ 10-8 per site per generation; recombination reshuffles them within meiosis; gene flow homogenises across populations at rate proportional to migration m; drift fluctuates frequencies at scale 1/√2N; selection moves them at rate ∼ sp(1-p). Real populations experience all five simultaneously, and their combined action is what drives observed evolution.
Hardy–Weinberg law; the five evolutionary forces are mutation, natural selection, genetic drift, gene flow and recombination.
Explain divergent evolution in detail. What is the driving force behind it?
Concept used. Divergent evolution is the process by which two or more groups of organisms descended from a common ancestor accumulate different heritable changes over time, ending up with markedly different forms. The classical morphological signature of divergent evolution is the existence of homologous organs –- organs with the same basic anatomical plan but adapted for different functions in different descendant species. The forelimbs of mammals (whale flipper, bat wing, cheetah leg, human arm) are the textbook example. The driving force is natural selection acting under different environmental pressures on geographically separated descendant populations (often accompanied by adaptive radiation). Mutation generates variation; environment-specific selection drives divergence; geographical isolation prevents gene flow that would otherwise homogenise the lineages.
- Start with a single ancestral population. Imagine an ancestral mammal ∼200 mya with a generalised five-fingered forelimb plan (humerus, radius–ulna, carpals, metacarpals, phalanges).
- Populations separate geographically. Continental drift, habitat fragmentation or migration leaves descendant populations in different environments –- one in water, one in the air, one on land.
- Selection pressures diverge. In water, fitness rewards broad flat paddles; in air, light bony wings; on land, long running legs. Each environment selects for different forelimb proportions and joint arrangements.
- Accumulated divergence. Over ∼100 myr, descendants with optimised forelimbs evolve: whale (paddle), bat (wing), cheetah (running leg), human (grasping arm). Bone-for-bone, their forelimbs match (homology); function-for-function, they differ.
- Other examples.
- Darwin's finches: one ancestral seed-eater on the Gal'apagos radiated into ∼13 species with different beak shapes for different foods (cactus, insects, large seeds, small seeds).
- Australian marsupials: a single ancestral marsupial diverged into kangaroo, koala, marsupial mole, Tasmanian wolf –- each filling a different ecological niche.
- Driving force. Natural selection under different environmental conditions imposed on geographically isolated sub-populations. Without environmental difference, selection would not drive divergence; without isolation, gene flow would re-mix the populations.
Divergent evolution: descendants of a common ancestor diverge into different forms by accumulating different heritable changes under different environmental selection pressures, producing homologous organs. Driving force: natural selection acting under different environmental conditions on geographically isolated descendant populations.
Strategic angle. Trace the ``why'' of divergence: same start, different environments, different selection, different outcomes.
- Common origin. Single ancestral population, generalised plan.
- Environmental diversification. Descendants face different habitats.
- Selection diversifies. Each habitat rewards a different body design.
- Outcome. Homologous organs across descendants –- identical bone plan, different functions.
Why this matters. Most large mammalian and bird groups today are the products of adaptive radiation (a form of divergent evolution) following ancestral isolation events.
Adaptive radiation as accelerated divergent evolution. Darwin's finches, Australian marsupials, Hawaiian honeycreepers and East African cichlids are textbook radiations –- a single ancestor in a niche-rich isolated setting produces dozens of descendant species in millions of years (or less). The driver is always natural selection acting on geographically separated descendants under different environmental challenges. Without isolation, gene flow would re-mix the populations and the same selection pressure would produce only one polymorphic species, not many distinct ones.
Mechanistic timeline. Step one is a single ancestral population in a uniform environment. Step two is a vicariance event (continental drift, river capture, mountain uplift, sea-level change) that splits it into geographically isolated descendant populations. Step three is the imposition of distinct local selection pressures; with no gene flow, the descendant populations cannot exchange alleles, so they drift apart genetically. Step four is the accumulation of reproductive isolation barriers (ecological, behavioural, gametic), at which point the descendants are full biological species. The whole cascade is divergent evolution, and natural selection is the engine that drives steps three and four.
Indian case study. The hangul deer of Kashmir and the swamp deer of Madhya Pradesh both descended from an ancestral Cervus form ∼2 Ma but diverged morphologically under different habitat pressures (cold mountain forest vs. warm wetland grass). Each retains the same basic cervid bone plan (homology), yet body size, antler architecture and coat density differ markedly. This is divergent evolution in action on Indian soil.
Divergent evolution = descendants of a common ancestor become different under different selection pressures, producing homologous organs. Driving force: natural selection acting under varying environments.
You have studied the story of Pepper moths in England. Had the industries been removed, what impact could it have on the moth population? Discuss.
Concept used. The Biston betularia peppered-moth story is the classic example of directional natural selection driven by human activity. Pre-Industrial Revolution: bark was pale (covered by lichens), pale moths were camouflaged from predatory birds, dark moths were eaten. Pale moths dominated. During the Industrial Revolution: soot killed lichens and blackened the bark; now dark moths were camouflaged and pale moths were eaten. The dark carbonaria form rose from <1% to >90% in heavily polluted cities like Manchester within ∼50 years. If the industries were removed, the environment would reverse: pollution drops, lichens recover, bark returns to pale –- and the selection pressure flips back.
- Restate the pre-removal state. Heavy industry ⇒ soot-blackened bark ⇒ selection favours dark (carbonaria) moths. Frequency of dark moths ∼95%.
- Industries removed: environmental recovery. Pollution falls; SO2 emissions drop; lichens regrow; bark surface gradually returns to its pale, lichen-coated, pre-industrial appearance over ∼10–30 years.
- Selection pressure flips. On pale bark, dark moths are once again conspicuous and eaten by birds; pale moths regain their camouflage advantage.
- Allele frequency response. The frequency of the dark carbonaria allele begins to fall; the frequency of the pale typica allele rises generation by generation. Real- world data after the 1956 UK Clean Air Act show exactly this reversal: dark-moth frequency in Manchester fell from ∼95% in 1959 to ∼10% by the 2000s.
- Time scale and rate. The reversal takes many generations –- roughly one moth generation per year. With strong selection differential, allele frequency can shift several percentage points per year, so a full reversal takes 30–50 years.
- Other possible outcomes.
- If the dark allele was very nearly fixed (close to 100%), the time for pale to recover is longer because pale individuals are rare initially.
- If pollution still lingers in patches, the population may settle at an intermediate equilibrium.
- The reversal is not fully complete in real data because residual urban heat, light pollution and surviving dark mutants keep dark frequency slightly above the pre-industrial baseline.
- Interpretation. The moth example confirms that natural selection acts continuously and reversibly –- whenever the environment changes, the population's allele frequencies track. It also illustrates how human action (industry, then clean-air policy) directly drives evolution in real time.
Removing the industries reverses the environment back to pale bark with lichens. Selection now favours pale (typica) moths again, so the dark (carbonaria) frequency falls and the pale frequency rises over several decades –- exactly what the UK Clean Air Act of 1956 produced in real data.
Strategic angle. Frame the moth population as a closed-loop feedback system: environment → selection → allele frequency → phenotype distribution. Flip the input, the output flips.
- Environment. Bark colour returns to pale.
- Selection. Direction reverses; pale favoured.
- Allele frequency. Dark falls, pale rises.
- Time. 30–50 yr for a near-complete reversal under realistic conditions.
Why this matters. The pepper-moth story is the canonical example that natural selection is contemporary, observable and human-modifiable –- a perfect demonstration for a NEET-level student.
Numerical prediction. If dark-morph fitness in pre-industrial conditions was ∼0.7 of pale-morph fitness, the dark frequency would fall by ∼3 percentage points per year under recovered clean-air conditions. Starting at ∼95% dark, a full reversal to below 5% dark would therefore take ∼30 years –- matching the observed 1956–2000 decline in Manchester moth records. The episode also doubles as the cleanest NEET example of allele-frequency tracking environmental change in real time.
Quantitative tracking. The 1956 UK Clean Air Act mandated coal-burning bans in declared smoke-control zones. Atmospheric SO2 in Manchester fell from ∼300 to ∼50 μg m-3 by 1980; Lecanora conizaeoides (an acid-tolerant lichen) returned, then Parmelia (more sensitive) by the 1990s. Concurrently, dark-morph Biston betularia frequency at Caldy Common fell from ∼95% in 1959 to ∼60% by 1985 to ∼10% by 2003 –- a textbook real-time allele-frequency shift recorded in field traps over 50 generations.
Connecting back to Hardy–Weinberg. The peppered-moth case is exactly a violation of the no-selection assumption: s for the dark allele under polluted bark is positive (selection for); under clean bark it flips to negative (selection against). The simple equation Δ p = sp(1-p) predicts the rate of change for each regime, and matches field data within experimental uncertainty.
The dark form would decline and the pale form would rise again over several decades, exactly mirroring the original industrial-era shift but in reverse.
What are the key concepts in the evolution theory of Darwin?
Concept used. Charles Darwin's theory of evolution by natural selection, set out in On the Origin of Species (1859), explains how populations of organisms change over generations and how new species arise. Darwin synthesised observations from his five-year voyage on HMS Beagle (particularly the Gal'apagos finches), Malthus's essay on population growth, animal-breeding practice and the geological work of Lyell. The theory rests on six interlocking key concepts.
- Key concept 1 –- Variation. Within any natural population, individuals differ from one another in heritable traits: size, colour, beak shape, fur length, behaviour. These variations arise spontaneously and are passed from parents to offspring.
- Key concept 2 –- Inheritance. Variations are heritable: parents pass their traits to offspring. (Darwin did not know about genes; that mechanism was Mendel's discovery, later integrated as the neo-Darwinian synthesis.)
- Key concept 3 –- Overproduction of offspring (struggle for existence). All species produce far more offspring than the environment can support. Malthus had shown that human populations grow geometrically while food supplies grow arithmetically; Darwin extended this to all organisms. The result is a continual ``struggle for existence'' –- competition for limited resources (food, mates, space).
- Key concept 4 –- Natural selection (survival of the fittest). In the struggle for existence, individuals with heritable variations that better suit them to the environment survive more often and leave more offspring. ``Fitness'' is measured by relative reproductive success, not strength. Spencer's phrase ``survival of the fittest'' captures this, with the understanding that ``fit'' means well-adapted, not just strong.
- Key concept 5 –- Origin of new species (descent with modification). Over very many generations, the accumulation of favoured variations transforms populations. If two sub-populations face different environments and are isolated, they diverge and eventually become reproductively incompatible –- giving rise to new species. All life therefore traces back to a common ancestor through a branching tree of descent with modification.
- Key concept 6 –- Gradualism. Evolutionary change is gradual, accumulating over thousands to millions of generations. Major novelties are not produced by single large jumps but by countless small modifications selected over time.
Darwin's six key concepts: (1) variation (individuals differ heritably); (2) inheritance (variations pass to offspring); (3) overproduction → struggle for existence (Malthusian); (4) natural selection (fittest leave more offspring); (5) descent with modification → origin of new species; (6) gradualism (slow cumulative change).
Strategic angle. Six pillars, building one on the next: variation → inheritance → struggle → selection → speciation → gradualism.
- Pillar 1. Heritable variation.
- Pillar 2. Overproduction creates competition.
- Pillar 3. Selection picks the better-adapted.
- Pillar 4. Cumulative change yields new species.
- Pillar 5. The process is gradual.
Why this matters. These six pillars unify every NEET evolution question –- finch beaks, peppered moths, antibiotic resistance, Hardy– Weinberg deviation are all worked examples of Darwin's six concepts.
Modern synthesis layer. What Darwin did not know –- and the 20th-century Modern Synthesis added –- is the molecular mechanism: Mendelian particulate inheritance (variations are not blended); mutation (de Vries, 1901) as the source of new variants; chromosomal segregation and recombination as the shufflers; and population-genetics mathematics (Hardy, Weinberg, Fisher, Haldane, Wright) as the bookkeeping. Darwin's six concepts remain intact; molecular biology only added the ``how'' behind each step, and that is the form of evolutionary theory NEET examines today.
Where Darwin succeeded and where he stalled. Darwin's six pillars (variation, inheritance, overproduction, selection, descent with modification, gradualism) explained what natural selection does and why adaptation arises, but he could not explain the mechanism of heredity, the source of new variants, or the discrete behaviour of genes. Each gap was plugged in turn by Mendel (particulate inheritance, 1866), de Vries (mutation, 1901), Sutton–Boveri (chromosomes, 1903), Watson–Crick (DNA, 1953) and Hardy–Weinberg/Fisher/Haldane/Wright (population-genetic mathematics, 1908–1940s). The combined edifice is the Modern Synthesis.
Variation, inheritance, struggle for existence, natural selection, descent with modification, gradualism.
Two organisms occupying a particular geographical area (say desert) show similar adaptive strategies. Taking examples, describe the phenomenon.
Concept used. When unrelated organisms living in the same environment independently evolve similar adaptive traits, the phenomenon is convergent evolution. The shared environment imposes shared selection pressures –- heat, water scarcity, low food –- so any lineage that solves these problems well is rewarded with survival and reproduction. Different lineages, starting from different ancestral body plans, end up with surprisingly similar solutions. The resulting structures are analogous organs: different ancestry, different anatomy, same function.
- Desert environment, selection pressures. High day temperature, intense solar radiation, very low and unpredictable rainfall, scarce food. To survive, any organism must minimise water loss, store water, regulate body temperature, and tolerate long fasts.
- Example 1 –- Desert plants. American cacti (family
Cactaceae) and African euphorbias (family Euphorbiaceae) are
unrelated angiosperm families. Yet both have evolved
independently:
- Succulent fleshy stems for water storage.
- Reduced or absent leaves (replaced by spines) to cut transpiration.
- Photosynthesis transferred from leaves to green stems.
- CAM photosynthesis (stomata open only at night).
- Spines for protection from herbivores.
- Example 2 –- Desert animals. The American kangaroo rat (rodent, placental mammal) and the Australian hopping mouse (rodent, also placental but on a different continent) and the African jerboa each independently evolved long hind legs for hopping locomotion, large ears for thermoregulation, the ability to produce highly concentrated urine to conserve water, and nocturnal habits. Different lineages, same desert solution.
- Example 3 –- Marsupial vs. placental. The Australian marsupial mole and the African placental golden mole, separated by 100 myr of evolution, look almost identical: cylindrical body, tiny eyes, powerful digging forelimbs, smooth fur. Both independently adapted to burrowing in sandy soil.
- Interpretation. Similar selection pressure + independent ancestry ⇒ analogous structures ⇒ convergent evolution. The phenomenon shows that the environment partly dictates form, regardless of starting genotype.
This phenomenon is convergent evolution: unrelated organisms (cacti vs. euphorbias; kangaroo rats vs. jerboas; marsupial vs. placental moles) independently evolve similar adaptive features under the same environmental selection pressures.
Strategic angle. Same environment, different ancestors, same solution = convergent.
- Pick the environment. Desert. Selection rewards water conservation.
- Pick the unrelated lineages. Cacti vs. euphorbias; or kangaroo rat vs. jerboa.
- List the shared adaptations. Succulent stem, spines, long hind limbs, concentrated urine, nocturnal habits.
Why this matters. Convergence is biology's strongest evidence that environment dictates form. The 2010s discovery of similar fish in isolated cave systems worldwide is a fresh example.
Edge case: desert birds. Sandgrouse (Africa, Asia) and the unrelated burrowing-owl (Americas) both evolved water-saving strategies in desert habitats: the sandgrouse soaks belly feathers in distant water and carries it back to chicks, the burrowing-owl uses subterranean cool air. Different lineages, different solutions –- but each solution converged on minimising water loss under desert selection. The same convergent logic explains why cacti, euphorbias and aloes (three unrelated angiosperm families) all evolved succulence and spines.
Plant convergence in detail. Cacti (family Cactaceae, Americas) and euphorbias (family Euphorbiaceae, Africa) shared a common angiosperm ancestor ∼120 Ma but have lived on separate continents for almost as long. Yet both independently lost their leaves (replaced by spines), thickened their stems into water-storage organs, shifted photosynthesis from leaves to stem chlorenchyma, adopted CAM (Crassulacean Acid Metabolism) photosynthesis to fix CO2 at night when stomata can open without losing water, and grew dense epidermal cuticles. Five independent biochemical and anatomical inventions converged in two unrelated families –- the most striking land-plant example of convergent evolution.
Why the environment is the architect. Selection rewards solutions that work in the local conditions. In a desert, any lineage that fails to conserve water is filtered out within a generation. The lineages that survive are the ones that –- by random heritable variation –- already had the seeds of water-saving traits. Over millions of years, these seeds are amplified into the succulent body plan we recognise, regardless of the lineage's starting morphology.
Convergent evolution; unrelated lineages independently arrive at similar adaptive solutions when they share the same environment.
We are told that evolution is a continuing phenomenon for all living things. Are humans also evolving? Justify your answer.
Concept used. Evolution is defined as change in allele frequencies in a population across generations. As long as any of the five evolutionary forces (mutation, natural selection, genetic drift, gene flow, recombination) is acting, evolution continues. Humans are biological organisms living in environments that present selection pressures (disease, diet, climate, mate choice), with active mutation, ongoing migration and finite population sub-structure –- so yes, humans are still evolving. The pace may be slower in some traits (cushioned by medicine, technology and culture) but is genuinely faster in others.
- Mutation is ongoing. Each newborn carries ∼70 new single-nucleotide mutations not present in either parent. These enter the human gene pool every generation. Mutation alone guarantees evolution.
- Natural selection is active. Documented examples in
living humans:
- Lactase persistence. Most adults worldwide lose the ability to digest lactose; in dairying populations of Europe and East Africa, mutations in LCT/MCM6 that keep lactase expressed into adulthood spread rapidly in the last ∼10 000 years.
- Sickle-cell allele. The HbS allele is maintained at high frequency in malarial regions because heterozygotes resist falciparum malaria –- a contemporary balanced polymorphism.
- High-altitude adaptation. Tibetans carry an EPAS1 variant that adjusts haemoglobin response to low oxygen; the allele rose to high frequency within the last few thousand years.
- Skin pigmentation. Variants in SLC24A5, SLC45A2, MC1R causing lighter skin spread in high-latitude populations after the migration out of Africa –- driven by vitamin-D selection.
- Gene flow is intense. Migration today is global. Genes flow between continents far faster than in any prior era. This is reshaping allele frequencies in real time.
- Genetic drift in subpopulations. Small isolated communities (some Amish, Faroese, Saudi tribal groups) show drift-driven allele-frequency shifts and elevated rare-disease frequencies –- ongoing evolution.
- Cultural buffering, not stopping. Modern medicine reduces the strength of some selection pressures (childhood diseases) but does not eliminate evolution. New pressures (antibiotic-resistant infections, dietary changes, environmental toxins) take their place. Mate choice (assortative mating by education, income, height) also creates new selection.
Yes, humans are still evolving. Active forces: continuous mutation (∼70 new mutations per newborn); documented natural selection on lactase persistence, sickle-cell, high-altitude adaptation and skin pigmentation; intense gene flow from global migration; genetic drift in isolated subpopulations. Evolution is a present-tense phenomenon in our species.
Strategic angle. Test each evolutionary force on the human population; if any is active, humans are evolving.
- Mutation. 70 new mutations per genome per generation. Active.
- Selection. Sickle-cell, lactase persistence, EPAS1, pigmentation alleles. Active.
- Gene flow. Globalisation drives intercontinental gene exchange. Active.
- Drift. Small isolated communities show drift signatures. Active.
Why this matters. Disease genetics, personalised medicine, and the GWAS revolution of the 2010s all rely on the fact that human variation is still being generated and filtered.
Single-generation cases. Even within the lifetime of a NEET aspirant reading this, allele frequencies of antibiotic-resistance genes in human-associated bacteria, CCR5-Δ32 (HIV resistance) in Eurasian populations and ALDH2 alcohol-tolerance in East Asian populations are measurably shifting. Evolution is not a museum exhibit –- it is the news. Modern genomic monitoring programmes (UK Biobank, Genome Asia 100K) catch these shifts as they happen, and the data feed directly into clinical genetics.
Genomic evidence today. The 1000 Genomes Project (2010s) catalogued ∼84 million SNPs across 26 human populations. Allele-frequency comparisons revealed dozens of loci under recent positive selection: EDAR (hair thickness in East Asians), TYRP1 and KITLG (skin pigmentation in Europeans), LCT (lactase persistence in Europeans and East Africans), HBB (sickle-cell trait in malarial Africa), HERC2 (eye colour in Europeans). Each is a measured allele-frequency shift over the last ∼10 000 years –- direct genomic proof that the human gene pool is still being remodelled by selection.
Yes, humans are evolving. All five evolutionary forces remain active in the human gene pool; specific allele frequencies are demonstrably shifting in living populations.
Had Darwin been aware of Mendel's work, would he been able to explain the origin of variations. Discuss.
Concept used. Darwin (1859) knew that variations occur and are heritable, but he did not know the mechanism of inheritance. He toyed with the wrong ``blending inheritance'' idea (parent traits average in offspring), which actually undermined his own theory –- blending would wipe out novel variations within a few generations. Gregor Mendel (1866) showed that inheritance is particulate: genes are discrete units that segregate cleanly in gametes and combine without diluting. Mendel's paper was published seven years after Origin of Species but ignored until 1900. Had Darwin known Mendel, he could have explained how variations persist across generations –- but the origin of new variations (mutation, recombination) was discovered even later. So the answer is: partly yes, partly no.
- What Darwin knew about variation. Variation exists; some is heritable; selection acts on heritable variation. But the source and mechanism of inheritance were a black box for him.
- Darwin's biggest problem: blending inheritance. The accepted view in Darwin's day was that offspring traits are averages of parent traits (like mixing paint). Under blending, any new beneficial variation would be halved every generation, diluting to nothing in ∼10 generations. Darwin worried about this openly in later editions of Origin.
- What Mendel solved. Particulate inheritance: traits are governed by discrete factors (later named genes) that segregate intact in gametes (Law of Segregation) and re-assort independently (Law of Independent Assortment). A new favourable variant does not blend; it retains its identity and can spread through the population unchanged in form.
- What Mendel did not explain. Mendel showed how existing variants are inherited, but he did not explain how new variants arise. The discovery of mutation (Hugo de Vries, 1901), chromosomal theory (Sutton, Boveri, 1903) and ultimately the structure of DNA (Watson, Crick, 1953) provided the modern answer.
- Putting it together: the Modern Synthesis (1930s–40s). Darwin (selection) + Mendel (particulate inheritance) + de Vries (mutation) + population genetics (Hardy, Weinberg, Fisher, Haldane, Wright) merged into the neo-Darwinian Modern Synthesis –- the framework biology still uses today.
- Answer to the question. If Darwin had known Mendel, he
could have:
- Defeated the blending-inheritance critique and saved his theory from its own internal worry.
- Explained why heritable variations persist across generations without dilution.
If Darwin had known Mendel: he would have explained the persistence and inheritance of variations (particulate inheritance defeats blending), saving his theory from the blending- inheritance critique. But the origin of new variations (mutation) needed the discoveries of de Vries (1901) and 20th-century molecular biology –- so Mendel alone would not have been enough.
Strategic angle. Split the question into two parts: persistence of variation (Mendel solves this) and origin of variation (Mendel does not).
- Persistence problem. Blending inheritance dilutes variants. Mendel's particulate inheritance fixes this –- alleles stay intact across generations.
- Origin problem. What creates new alleles? Mendel does not say. The answer is mutation (de Vries, 1901) and recombination, only fully understood with the DNA structure (Watson–Crick, 1953).
- Synthesis. Darwin + Mendel + de Vries = neo-Darwinian Modern Synthesis.
Why this matters. The history of biology is the story of patching one gap (variation source) to plug another (variation persistence). Knowing both gaps and both patches answers any neo-Darwinian NEET question.
Resolution of the question. Mendel could have rescued Darwin's theory from its own internal blending-inheritance critique, because particulate inheritance preserves new variants undiluted across generations. But Mendel did not explain where new alleles come from –- that gap required Hugo de Vries's mutation theory (1901), Sutton–Boveri chromosome theory (1903) and finally the Watson–Crick DNA structure (1953). So if Darwin had known Mendel, the theory would have become much stronger on the inheritance side, but its account of variation's origin would have remained incomplete until 20th-century molecular biology.
Historical irony. Darwin owned a copy of the journal in which Mendel's 1866 paper was published, but never cut the pages of Mendel's article –- he had received it from a colleague but did not read it. Had he done so, the Modern Synthesis might have arrived 70 years earlier and the early 20th-century ``mutationist vs. Darwinist'' debate would have been short-circuited. The episode is a sobering reminder that the right idea reaching the right reader at the right time is itself a contingent historical event.
Mendel would have explained how variations persist (particulate inheritance defeats blending). The origin of new variations required mutation (de Vries, 1901) –- a discovery later than both Darwin and Mendel.
NCERT Exemplar Solutions for Class 12 Biology: All Chapters
| Chapter | Exemplar Solutions Link |
|---|---|
| Chapter 1 | Sexual Reproduction in Flowering Plants Exemplar Solutions |
| Chapter 2 | Human Reproduction Exemplar Solutions |
| Chapter 3 | Reproductive Health Exemplar Solutions |
| Chapter 4 | Principles of Inheritance and Variation Exemplar Solutions |
| Chapter 5 | Molecular Basis of Inheritance Exemplar Solutions |
| Chapter 7 | Human Health and Disease Exemplar Solutions |
| Chapter 8 | Microbes in Human Welfare Exemplar Solutions |
| Chapter 9 | Biotechnology Principles and Processes Exemplar Solutions |
| Chapter 10 | Biotechnology and Its Applications Exemplar Solutions |
| Chapter 11 | Organisms and Populations Exemplar Solutions |
| Chapter 12 | Ecosystem Exemplar Solutions |
| Chapter 13 | Biodiversity and Conservation Exemplar Solutions |
Frequently Asked Questions on Evolution Class 12 Biology Exemplar Solutions
How many problems does the NCERT Exemplar Class 12 Biology Chapter 6 contain?
The Exemplar carries 52 problems split across 18 MCQ items, 15 Very Short Answer (VSA), 12 Short Answer (SA), and 7 Long Answer (LA) questions, every one of them answered in this Collegedunia PDF with full reasoning and an Expert's Solution.
Are the Class 12 Biology Chapter 6 Evolution Exemplar Solutions enough for NEET?
Yes for recall and phrasing, no for full coverage. The Exemplar locks the high-yield NEET phrases (Hardy-Weinberg 2pq, adaptive radiation, industrial melanism, brain-volume timeline), but NEET aspirants should also pair it with the previous-year question set for assertion-reason items.
Is Evolution still part of the 2026-27 NCERT syllabus for Class 12 Biology?
Yes. The current 2026-27 NCERT retains Chapter 6 Evolution in full, including origin of life, evidences of evolution, Hardy-Weinberg, adaptive radiation, and human evolution. No sub-topic was dropped, so every Exemplar problem on this page is examinable.
Which is the most asked Exemplar question type in Evolution?
MCQ items dominate by count, 18 of the 52 questions, and they map directly onto NEET's single-correct format. Within MCQ, Hardy-Weinberg numericals and the homologous-versus-analogous pair are the two highest-frequency topics.
How is the Exemplar harder than the NCERT textbook for Chapter 6 Evolution?
The textbook asks "state" and "define", the Exemplar asks "calculate" and "differentiate". For example, NCERT asks the Hardy-Weinberg principle, the Exemplar asks you to compute 2pq for a given q. The step-up is from recall to numerical mechanism, which is exactly what NEET expects.
Can I download the Evolution Exemplar Solutions PDF for free?
Yes, the full PDF is free to download from the card above. It covers all 52 problems, includes the Expert's Solution after every question, and is mapped to the 2026-27 NCERT chapter for Class 12 Biology Chapter 6 Evolution.
What are the most common mistakes students make in Evolution Exemplar questions?
Confusing analogous and homologous organs, using q2 instead of 2pq for heterozygotes, mixing up the order from Australopithecus to Homo sapiens, defining natural selection as "survival of the fittest" without mentioning differential reproduction, and confusing Lamarckism with Darwinism. All five mistakes are corrected inside the PDF.


Comments